alteraciones neuropsicolÓgicas en consumidores de cocaÍna …hera.ugr.es/tesisugr/21168581.pdf ·...
TRANSCRIPT

Laura Moreno López
ALTERACIONES NEUROPSICOLÓGICAS EN CONSUMIDORES DE COCAÍNA:
CORRELATOS NEUROANATÓMICOS

UNIVERSIDAD DE GRANADA
Departamento de Personalidad, Evaluación y Tratamiento Psicológico
TESIS DOCTORAL
ALTERACIONES NEUROPSICOLÓGICAS EN CONSUMIDORES DE COCAÍNA:
CORRELATOS NEUROANATÓMICOS
Doctoranda: Laura Moreno López
Directores: Dr. Antonio Javier Verdejo García y Dr. Miguel Pérez García

Editor: Editorial de la Universidad de GranadaAutor: Laura Moreno LópezD.L.: GR 1203-2013ISBN: 978-84-9028-529-9

Los directores Dr. Antonio Javier Verdejo García y Dr. Miguel Pérez García
autorizan la presentación de la tesis doctoral titulada: “Alteraciones Neuropsicológicas en
consumidores de cocaína: Correlatos neuroanatómicos” presentada por Dña. Laura
Moreno López.
Fdo. Dr. Antonio Javier Verdejo García Fdo. Dr. Miguel Pérez García
Fdo. Laura Moreno López

A mis padres

he future belongs to those who believe in the beauty of their dreams
(Eleanor Roosevelt)
T

Agradecimientos
Quiero agradecer la realización de este trabajo a todos aquellos que lo han
hecho posible y muy especialmente a mis tutores Antonio Verdejo García y Miguel
Pérez García, por su apoyo y animo constantes y su ayuda incondicional.
A Emmanuel Stamatakis y Maki Kasahara por estar conmigo en los buenos y
malos momentos de mi primera estancia y por ayudarme en todo lo que necesité y he
necesitado hasta el día de hoy. Siempre seréis lo mejor de Cambridge para mí.
A Rita Goldstein, Nelly Alia-Klein, Tom Maloney, Patricia Woicik, Anna Konova
y Muhammad Parvaz del Departamento de Medicina del laboratorio Brookhaven de
Nueva York y a Martin Paulus y Scott Mackey del Departamento de Psiquiatría de la
Universidad de California San Diego, por darme la oportunidad de trabajar con ellos y
aprender un poquito más de este apasionante campo.
Asimismo, no quiero olvidarme de Carles Soriano Más por su apoyo y su ayuda
constantes, de todos mis compañeros del grupo de Neuropsicología y
Neuroinmunología clínica y de todos los participantes voluntarios de mis estudios.
Y por último, y en mayúsculas, a aquellos sin los que nada de esto tendría
sentido, mis padres, mi hermano y Jorge.

[ÍNDICE]

Presentación 1 I. INTRODUCCIÓN 5 1. Introducción 6 2. Exploración de los factores neurobiológicos y neuropsicológicos implicados en la 8 adicción: Neuroimagen y neuropsicología de la impulsividad y las funciones ejecutivas
2.1. Neuroimagen y adicciones 8
2.1.1. Clasificación y características de las técnicas de neuroimagen 9
2.1.1.1. Resonancia magnética estructural 9 2.1.1.2. Tomografía por emisión de positrones 10
2.1.2. Hallazgos de neuroimagen en consumidores de distintas drogas 11
2.1.2.1. Cocaína 12 2.1.2.2. Heroína 13 2.1.2.3. Alcohol 14 2.1.2.4. Éxtasis (MDMA) 16 2.1.2.5. Cannabis 17
2.2. Impulsividad y adicciones 18
2.2.1. Instrumentos de evaluación de la impulsividad 19 2.2.1.1. Inventarios de personalidad impulsiva 19 2.2.1.2. Medidas de control inhibitorio 20 2.2.2. Hallazgos relacionados con la impulsividad en consumidores 21 de distintas drogas 2.2.2.1. Cocaína 21 2.2.2.2. Heroína 23 2.2.2.3. Alcohol 23 2.2.2.4. Éxtasis (MDMA) 24 2.2.2.5. Cannabis 25 2.3. Funciones ejecutivas y adicciones 25 2.3.1. Evaluación neuropsicológica de las funciones ejecutivas 26 2.3.1.1. Actualización 27 2.3.1.2. Control inhibitorio 28 2.3.1.3. Flexibilidad cognitiva 28 2.3.1.4. Toma de decisiones 28 2.3.1.5. Procesos emocionales 29 2.3.2. Hallazgos neuropsicológicos relacionados con las funciones 30 ejecutivas en consumidores de distintas drogas 2.3.2.1. Cocaína 31 2.3.2.2. Heroína 32 2.3.2.3. Alcohol 32 2.3.2.4. Éxtasis (MDMA) 33 2.3.2.5. Cannabis 34

II. JUSTIFICACIÓN Y OBJETIVOS 35 III. MEMORIA DE TRABAJOS 41 Artículo 1. Trait impulsivity and prefrontal gray matter reductions in cocaine 43 dependent individuals 1. Introduction 44 2. Materials and methods 46
2.1. Participants 46 2.2. Instruments and assessment procedures 47 2.2.1. Patterns of drug use 47 2.2.2. Trait impulsivity 49 2.3. MRI adquisition 49 2.4. Image analysis 50 2.4.1. Global effects of patterns of drug use 50 2.4.2. Regional GM and WM differences between cocaine dependent 51 individuals and non-drug using controls 2.4.3. Regional relationships between GM and WM and measures of 51 impulsivity and estimates of drug use 3. Results 51
3.1. Participants’ characteristics 51 3.2. Trait impulsivity 52 3.3. Imaging analysis 52 3.3.1. Global effects 52 3.3.2. Regional GM and WM differences between cocaine dependent 53 individuals and non-drug using controls 3.3.3. Regional relationships between GM and WM and measures of 56 impulsivity and estimates of drug use 4. Discussion 58 References 62 Artículo 2: Neural Correlates of the Severity of Cocaine, Heroin, Alcohol, 67 MDMA and Cannabis Use in Polysubstance Abusers: A Resting-PET Brain Metabolism Study 1. Introduction 68 2. Methods 69
2.1 Participants 69
2.2 Testing protocols and procedures 70 2.3 Tools 70
2.3.1 Drug Use Information 70
2.3.2 PET Image Acquisition 72
2.4 Data analysis 72
2.4.1 Preprocessing of PET images 72
2.4.2 Statistical Analysis 72

3. Results 73
3.1 Amount of use 73
3.2 Duration of use 74
4. Discussion 77 References 81 Artículo 3: Neural correlates of hot and cold executive functions in 85 polysubstance addiction: Association between neuropsychological performance and resting-PET brain metabolism 1. Introduction 86 2. Methods 88
2.1. Participants 88
2.2. Testing protocols and procedures 90
2.3. Instruments 90
2.3.1. Patterns of drug use 90
2.3.2. Neuropsychological tests 91
2.3.3. PET image acquisition 92
2.4. Data analysis 92
2.4.1. Preprocessing of PET images 92
2.4.2. ROI analysis 92
2.4.3. Statistical analyses 93
2.4.3.1. Behavioral analysis 93
2.4.3.2. ROI analysis 93
2.4.3.3. Voxel based analysis 93
3. Results 94
3.1. Neuropsychological performance 94
3.2. ROI analyses 95
3.2.1. Association between neuropsychological performance and ROIs 95
3.2.2. Association between the ROIs associated with 95 neuropsychological performance and estimates of drug use
3.3. Voxel-based analyses 96
3.3.1. Cold executive functions –whole brain voxel-based correlates 98 in SDI
3.3.2. Hot executive functions –whole brain voxel-based correlates 100 in SDI
3.3.3. The relationship between voxel-based derived regions 102 associated with neuropsychological performance and estimates of drug use
4. Discussion 102 References 107 Supplementary material 116

IV. DISCUSIÓN 121 1. Discusión general 122 2. Conclusiones 128 3. Perspectivas de futuro 129 DOCTORADO EUROPEO 131 1. Summary 132 2. Conclusions 136 3. Future perspectives 137 REFERENCIAS 139 ANEXOS 171

P á g i n a | 1
PRESENTACIÓN [ ]

P á g i n a | 2
El informe mundial sobre drogas emitido por Naciones Unidas en 2011 (UNODC,
2011) estimó que en 2009, entre 149 y 272 millones de personas con edades
comprendidas entre los 15 y los 64 años habría consumido drogas ilegales en al menos
una ocasión durante el último año, al menos la mitad lo habría hecho durante el último
mes y entre 15 y 39 millones de personas tendrían problemas con ese consumo.
A nivel mundial, el cannabis es la droga ilegal más utilizada seguida por las
anfetaminas, los opiáceos y la cocaína. Sin embargo, el patrón de consumo es diferente en
Europa, donde la cocaína es la segunda droga ilegal más consumida seguida por las
anfetaminas, el éxtasis y los opiáceos. La prevalencia del consumo de cocaína difiere entre
los países que conforman la Unión Europea pero es especialmente problemática entre
varones jóvenes de entre 15 y 34 años y en países como Dinamarca, Irlanda, Italia, España
y Reino Unido. Y es que, aunque la prevalencia del consumo es menor en la Unión Europea
(2,1%) que en el mismo colectivo en Australia (3,4%), Canadá (3,3%) o Estados Unidos
(4,1%), en España (4,4%) y Reino Unido (4,8%) ha presentado cifras más elevadas
(EMCDDA, 2011).
Aunque el consumo de drogas ilegales constituye uno de los mayores problemas de
nuestra sociedad, no debemos olvidar el efecto que el abuso o co-abuso de otras sustancias
legales como el alcohol tiene en la población. Según la organización mundial de la salud, el
alcoholismo es una de las drogodependencias más extendidas a nivel mundial y constituye
el tercer lugar entre los principales factores de riesgo de muerte prematura y discapacidad
en el mundo. En nuestra sociedad, el consumo de alcohol es una práctica profundamente
arraigada y se estima que la mayoría de la población consume bebidas alcohólicas
esporádica o habitualmente (OED, 2009). Según el último informe del Observatorio
Español sobre Drogas, el consumo de alcohol parece haberse establecido e incluso ha
descendido en algunos sectores de la población, pero en los últimos años, el consumo
entre los jóvenes, un consumo caracterizado por tomar grandes cantidades de alcohol

P á g i n a | 3
durante breves periodos de tiempo (el denominado binge drinking), que ha empezado a
asociarse con la presencia de alteraciones neuropsicológicas en estas poblaciones (Parada
et al., 2012; Squeglia et al., 2012), está alcanzando cifras alarmantes.
Además de sus evidentes repercusiones sobre la salud física y psicológica de los
consumidores, el consumo de drogas tiene importantes implicaciones familiares, sociales,
culturales, políticas y económicas, por lo que constituye un fenómeno de enorme
complejidad, pero también de relevancia equiparable. Como consecuencia de su
complejidad, este fenómeno puede ser abordado desde múltiples perspectivas científicas.
En este sentido, diversas fuentes de evidencia, incluyendo estudios preclínicos en
animales, estudios farmacológicos, neuropsicológicos y de neuroimagen cerebral, han
destacado la relevancia de las alteraciones cognitivas y emocionales encontradas en
individuos drogodependientes o con alto riesgo de iniciarse en el consumo. Especialmente,
ha aumento el interés por conocer los correlatos neuropsicológicos y neuroanatómicos del
consumo de distintas drogas, así como la importancia que ciertos rasgos de personalidad
como la impulsividad pueden tener en el inicio, la progresión del consumo recreativo a la
dependencia y la recaída en estos pacientes (Verdejo-García, Lawrence, & Clark, 2008).
En los últimos años, el extraordinario desarrollo de las técnicas de neuroimagen
cerebral ha permitido investigar con mayor precisión la naturaleza, localización y
extensión de dichas alteraciones y en muchas ocasiones, se ha mostrado como la
herramienta más eficaz para detectar alteraciones a nivel cerebral en consumidores de
ciertas sustancias psicoactivas (p.e. Barrós-Loscertales et al., 2011a; Tapert et al., 2007).
Sin embargo, la presencia de ciertas limitaciones inherentes al estudio del fenómeno de la
drogadicción ha llevado a resultados inconsistentes y en muchas ocasiones
contradictorios.
En el contexto de aportar conocimientos sobre esta temática y superar las
limitaciones existentes en estudios previos de adicción, la presente Tesis Doctoral tuvo

P á g i n a | 4
como objetivo general el estudio de las alteraciones cerebrales estructurales y funcionales
presentes en individuos policonsumidores de drogas y (i) las variables de personalidad
que incrementan la predisposición al consumo y la dependencia, (ii) las estimaciones de
cantidad y duración de consumo de diferentes drogas, y (iii) el funcionamiento
neuropsicológico de los procesos ejecutivos típicamente afectados por el consumo de
drogas.

P á g i n a | 5
INTRODUCCIÓN [ ]

P á g i n a | 6
1. Introducción
Durante la primera mitad del siglo XX, la adicción fue despreciada en su carácter de
enfermedad, siendo considerada fundamentalmente un problema de índole moral o
antisocial. Los modelos clásicos de la adicción habían enfatizado el papel del llamado
“circuito de la recompensa” o “del placer” (el circuito mesolímbico dopaminérgico) y se
defendía que las drogas se consumían porque eran reforzadores potentes y, por tanto,
placenteras. Los trastornos adictivos eran clasificados en las primeras ediciones del
Manual Diagnóstico y Estadístico de los Trastornos Mentales (DSM) y la Clasificación
Internacional de Enfermedades (CIE) como trastornos de la personalidad o “trastornos de
carácter, de conducta y de inteligencia”.
Efectivamente, los primeros contactos con las drogas producen efectos placenteros
en las personas que las consumen y durante mucho tiempo, estos efectos han sido
asociados con la activación del circuito mesolímbico dopaminérgico. Utilizando los
mismos mecanismos fisiológicos que los reforzadores naturales, las drogas actúan sobre
este sistema pero a distintos niveles (Nestler, 2005). Así, la cocaína, las anfetaminas y la
nicotina producirían la liberación de dopamina en el núcleo accumbens, los opiáceos
producirían la activación de los receptores de péptidos opioides en el área tegmental
ventral y el núcleo accumbens y el alcohol produciría la activación de sistemas GABA en el
núcleo accumbens y la amígdala.
Sin embargo, a medida que avanza el consumo de la sustancia, se producen un
conjunto de neuroadaptaciones en las regiones asociadas con los efectos reforzantes de las
drogas y en aquellas conectas neuroanatomicamente con estas que conllevan el paso del
consumo de drogas por sus efectos reforzantes, al consumo de estas de forma impulsiva y
finalmente compulsiva, el consumo crónico y la recaída. Esta transición implica la
reprogramación de circuitos neuronales implicados entre otros en el refuerzo y la
motivación, la memoria, el funcionamiento ejecutivo y la regulación emocional. De entre

P á g i n a | 7
los diferentes circuitos implicados, los más estudiados en los últimos años se localizan en
las áreas prefrontales y se han propuesto como bases anatómicas del control de impulsos,
la toma de decisiones y la regulación emocional (Goldstein & Volkow, 2002, 2011).
En la actualidad, la adicción se define como un trastorno crónico y recidivante
caracterizado por un consumo de drogas abusivo y persistente a pesar de sus crecientes
consecuencias negativas para la vida de la persona (DSM-IV). La clasificación CIE-10
introduce el matiz de que la adicción tiene un carácter compulsivo y se caracteriza por la
falta de control por parte del individuo.
Definidas de esta manera, las alteraciones que caracterizan la adicción tienen
importantes correlatos neuropsicológicos al nivel de una presumible disfunción de las
habilidades encargadas de organizar y programar conductas dirigidas a objetivos y tomar
decisiones adaptativas. En este sentido, las alteraciones neuropsicológicas pueden
contribuir significativamente al consumo y la adicción a través de al menos dos
mecanismos (Rogers & Robbins, 2001). En primer lugar, la presencia de alteraciones
neuropsicológicas puede incrementar la probabilidad de conductas de búsqueda y
consumo de drogas tanto en las fases iniciales del consumo como en las recaídas y facilitar
el paso de un consumo recreativo a la dependencia (George & Koob, 2010; Verdejo-García
et al., 2008). En segundo lugar, la existencia de déficits neuropsicológicos puede limitar o
interferir la capacidad de los individuos drogodependientes para asimilar los contenidos y
las actividades de los programas de rehabilitación (Aharonovich, Amrhein, Bisaga, Nunes,
& Hasin, 2008; Streeter et al., 2008; Turner, LaRowe, Horner, Herron, & Malcolm, 2009).
De acuerdo con su relevancia clínica y teórica, el estudio de las alteraciones
neuropsicológicas asociadas al consumo de drogas y su relación con los mecanismos
cerebrales implicados en la adicción ha experimentado un considerable avance en los
últimos años, con importantes aportaciones empíricas derivadas de modelos animales,
estudios farmacológicos y estudios de neuroimagen cerebral. El desarrollo y la aplicación

P á g i n a | 8
de las técnicas de neuroimagen se ha revelado como una potente herramienta de carácter
transversal (que puede emplearse en estudios con animales, y en estudios de farmacología
clínica, neuropsicología y tratamiento en humanos) para mejorar nuestra comprensión de
los procesos adictivos.
2. Exploración de los factores neurobiológicos y neuropsicológicos implicados en la
adicción: Neuroimagen y neuropsicología de la impulsividad y las funciones
ejecutivas
A continuación se presenta una breve revisión de las principales modalidades de
exploración neurobiológica y neurocognitiva en el estudio de los procesos de impulsividad
y funcionamiento ejecutivo en el campo de las adicciones. En cada apartado se presentará,
en primer lugar, un breve resumen de las principales técnicas utilizadas y, en segundo
lugar, un resumen de los principales hallazgos resultantes de la aplicación de las mismas
en consumidores de cocaína, heroína, alcohol, MDMA y cannabis (las drogas que
presentaron una mayor prevalencia de consumo entre los participantes de nuestros
estudios). Nos centraremos en los resultados del consumo crónico de las sustancias,
evitando los estudios de sus efectos agudos o de aquellos estudios que se han llevado a
cabo tras breves periodos de abstinencia (<48h), pues son las alteraciones a largo plazo las
que tienen un impacto más directo y significativo sobre la rehabilitación y el
funcionamiento diario de los individuos drogodependientes, incluso una vez abandonado
el consumo de drogas (Verdejo-García, López-Torrecillas, Giménez, & Pérez-García, 2004).
2.1. Neuroimagen y adicciones
Las técnicas de imagen cerebral se han convertido en un pilar fundamental para el
desarrollo de las neurociencias, permitido en los últimos años examinar “in vivo” los
efectos agudos de la administración de sustancias psicoactivas, los correlatos cerebrales
del deseo intenso por consumir las drogas (craving), o las alteraciones a largo plazo en
regiones cerebrales y sistemas neuroquímicos implicados en el consumo crónico y la

P á g i n a | 9
dependencia. Desde los inicios de su aplicación, y en un breve periodo de tiempo, los
hallazgos de neuroimagen cerebral han contribuido de manera significativa a comprender
los sustratos cerebrales de las drogodependencias y sus repercusiones sobre el
funcionamiento neuropsicológico de los consumidores.
2.1.1. Clasificación y características de las técnicas de neuroimagen
En función de su aplicación, las técnicas de neuroimagen pueden ser agrupadas en
dos grandes categorías: técnicas estructurales y técnicas funcionales. Las técnicas
estructurales informan sobre la localización, forma y tamaño de algunas regiones
cerebrales y permiten cuantificar los cambios volumétricos o de densidad de la sustancia
gris y la sustancia blanca cerebral. Las técnicas funcionales miden los cambios en la
actividad, el metabolismo cerebral o en ciertos parámetros neurofarmacológicos como la
densidad de receptores o los niveles de neurotransmisores y metabolitos.
Las técnicas estructurales más utilizadas en la investigación en drogodependencias
son la resonancia magnética estructural y las imágenes por tensor de difusión. Las técnicas
funcionales más utilizadas son la resonancia magnética funcional, la tomografía por
emisión de positrones (PET) y la tomografía por emisión de fotón único (SPECT). Aunque
el coste del SPECT es menor, el uso de la tomografía por emisión de positrones presenta
importantes ventajas con respecto a este, entre las que cabe destacar una localización más
precisa del trazador y una mejor resolución espacial.
2.1.1.1. Resonancia magnética estructural
Para que podamos obtener una imagen del interior de un objeto, sin que esto
conlleve la rotura o descomposición de su estructura interna, es necesario hacer llegar
hasta él ondas con forma conocida, ya sea por reflejo (ultrasonidos), por absorción y
transmisión o por emisión desde el interior, e intentar captar la modificación que se ha
producido en esa onda ya sea en su fase, en su frecuencia o en su amplitud. El tipo de

P á g i n a | 1 0
ondas que se emplean en resonancia son ondas electromagnéticas a frecuencias de radio
del orden de los megahertzios y los receptores y posteriormente los emisores del interior
del cuerpo humano son los protones de algunos núcleos atómicos que hacen de antena
emisora y receptora. El escáner de resonancia magnética consta de un campo magnético
muy intenso, homogéneo y uniforme. Cuando situamos a la persona bajo la influencia de
ese campo, los protones de sus tejidos se alinean con respecto a este y cuando dicha
energía cesa, el núcleo que ha captado esa energía la devuelve, y esta puede ser captada
desde el exterior mediante un receptor de campo magnético adecuado. Esta información
es empleada a continuación para construir una imagen con un alto nivel anatómico que
puede ser visualizada en dos o tres dimensiones y que tras ser analizada permitirá la
determinación del volumen, la densidad, la localización y la composición de la materia gris
y materia blanca cerebral.
2.1.1.2. Tomografía por emisión de positrones
La tomografía por emisión de positrones es una técnica no invasiva de diagnóstico
por imagen que permite determinar el nivel de actividad metabólica de los diferentes
tejidos del cuerpo humano, especialmente del sistema nervioso central, tras la
administración de una sustancia marcada radiactivamente. Como norma general se marca
alguna sustancia química como el oxígeno, el hidrógeno o la glucosa que será inyectada en
la sangre del paciente. Una vez fijada al tejido y transcurrido un tiempo (variable para cada
sustancia radioactiva), los átomos inestables del isótopo liberarán positrones que se
aniquilarán al contactar con los electrones de otros átomos circundantes. Dicho proceso de
aniquilación generará dos fotones que se desplazarán a la misma velocidad pero en
sentido opuesto y que serán detectados por un tomógrafo externo o sistema de detección
que mediante detectores de fotones situados en forma de cilindro alrededor de la cabeza,
será capaz de mapear el origen del proceso de aniquilación protón-electrón, y por tanto,
estimar la localización del proceso metabólico de interés (Mishina, 2008) (Figura 1).

P á g i n a | 1 1
Figura 1. Esquema del proceso de captura de la tomografía por emisión de positrones
De entre los diferentes radiofármacos emisores de positrones, el más utilizado es
el Flúor-18, un radiofármaco que al unirse a la 2-O-trifluorometilsulfonil manosa permite
la obtención del trazador 18-Flúor-Desoxi-Glucosa (18-FDG). La administración de este
radioisótopo permitirá identificar, localizar y cuantificar el consumo de glucosa de las
diferentes células del cerebro, permitiéndonos conocer que áreas se encuentran
hiperactivadas y cuales hipoactivadas. El uso de 18-FDG ha mostrado su validez en la
evaluación de los procesos adictivos, permitiéndonos conocer los efectos de la
administración aguda de una droga (Volkow et al., 2008), la respuesta a la presentación de
estímulos condicionados con esta (Volkow et al., 2011), el craving (Wang et al., 1999) o el
resultado de su consumo crónico (Galynker et al., 2007).
2.1.2. Hallazgos de neuroimagen en consumidores de distintas drogas
En esta sección presentamos los principales hallazgos derivados de la aplicación de
la resonancia magnética estructural y la tomografía por emisión de positrones, las técnicas
Unidad de Procesamiento
Aniquilación Reconstrucción de las imágenes

P á g i n a | 1 2
de neuroimagen cerebral utilizadas en nuestros estudios, a la investigación de los efectos
crónicos del consumo de cocaína, heroína, alcohol, MDMA y cannabis sobre la estructura o
morfología cerebral y sobre el metabolismo cerebral de la glucosa.
2.1.2.1. Cocaína
La cocaína produce sus efectos estimulantes a través de la inhibición de la
recaptación de los neurotransmisores dopamina, serotonina y norepinefrina. Se ha
propuesto que las alteraciones neuropsicológicas y de neuroimagen vinculadas al
consumo de cocaína están asociadas a neuroadaptaciones provocadas por la sobre-
estimulación de las vías dopaminérgicas y la consecuente hipo-activación o el agotamiento
de estas vías una vez abandonado el consumo (Gruber & Yergelun-Todd, 2001).
Alteraciones estructurales
La mayoría de los estudios que se han llevado a cabo con esta población han
encontrado reducciones del volumen de materia gris y materia blanca cerebral en las
cortezas prefrontal dorsolateral y orbitofrontal, frontal y temporal y en estructuras
subcorticales como la ínsula y en el cerebelo (Barrós-Loscertales et al., 2011b; Franklin et
al., 2002; Makris et al., 2004b; Matochik, London, Eldreth, Cadet, & Bolla, 2003; Sim et al.,
2007; Tanabe et al., 2009).
Alteraciones funcionales
La mayoría de los estudios de neuroimagen cerebral llevados a cabo en este campo
se han dirigido al estudio de la alteración de las vías dopaminérgicas (Martinez &
Narendran, 2010). Los pocos estudios que han evaluado el funcionamiento cerebral en
situación de reposo con PET han observado reducciones del metabolismo cerebral
preferentemente en áreas prefrontales (p.e. corteza prefrontal dorsolateral, orbitofrontal
y corteza anterior del cíngulo). En uno de los primeros estudios dirigidos a estudiar los
efectos a largo plazo del consumo de drogas en el metabolismo cerebral de consumidores

P á g i n a | 1 3
de cocaína en situación de abstinencia, Volkow y colaboradores (1992) encontraron
reducciones significativas de la actividad metabólica en la corteza frontal que persistieron
tras varios meses de abstinencia. Además, las alteraciones encontradas correlacionaron
con la dosis y la duración del consumo de cocaína. En otro estudio del mismo grupo, se
encontraron reducciones significativas del metabolismo regional del córtex prefrontal
(relacionadas con una menor concentración de receptores de dopamina en esta región) en
consumidores de cocaína (Volkow et al., 1993).
2.1.2.2. Heroína
La heroína y otros opiáceos ejercen sus efectos cerebrales a través de su acción
sobre los receptores específicos mu, delta y kappa, que se expresan en diversas áreas
relacionadas con los efectos reforzantes de las drogas. Estas sustancias también
incrementan la producción de dopamina, pero a través de una vía indirecta, al reducir la
actividad inhibitoria del GABA en el área tegmental ventral (Camí & Farré, 2003).
Muchos de los estudios encontrados en esta población presentan importantes
limitaciones metodológicas, entre las que cabe destacar el uso de pacientes en tratamiento
con metadona o de pacientes con importantes niveles de co-abuso de otras drogas (Lyoo
et al., 2004; Pezawas et al., 2002; Reid et al., 2008).
Alteraciones estructurales
Los estudios llevados a cabo en consumidores de heroína en situación de
abstinencia han mostrado discrepancias importantes. No obstante, en los últimos años, el
desarrollo de nuevas técnicas y análisis de neuroimagen cerebral ha permitido demostrar
la existencia de reducciones significativas de la materia gris cerebral en regiones
prefrontales y temporales en estas poblaciones (Pezawas et al., 1998; Yuan et al., 2009).
Pezawas y colaboradores (1998) encontraron ensanchamientos del espacio ventricular y
pericortical en consumidores de heroína en situación de abstinencia, revelando

P á g i n a | 1 4
reducciones del volumen cerebral en estos individuos. Asimismo, el volumen regional del
córtex frontal estaba correlacionado con la duración de la abstinencia en los consumidores
de heroína, de modo que la pérdida de volumen frontal era significativamente inferior en
aquellos individuos con periodos de abstinencia superiores a un año. En el estudio llevado
a cabo por Yuan y colaboradores (2009), se encontró una reducción significativa de la
densidad de materia gris cerebral en la corteza prefrontal, temporal y el cíngulo y análisis
posteriores controlando el efecto de la edad, el nivel educativo, el género, el uso de
nicotina y la duración de la abstinencia mostraron que la duración del consumo de heroína
correlacionaba negativamente con la densidad de la materia gris de estos pacientes.
Alteraciones funcionales
Los estudios del metabolismo cerebral llevados a cabo en estas poblaciones con
PET también son contradictorios. En este sentido, mientras que Galynker y colaboradores
(2000) encontraron aumentos significativos del metabolismo cerebral en la parte anterior
del cíngulo en consumidores de heroína en situación de abstinencia que habían recibido
tratamiento con metadona durante varios años, un estudio de este mismo grupo encontró
que los consumidores de opiáceos mostraban reducciones significativas del metabolismo
cerebral en las partes medial y anterior del cíngulo, ínsula y corteza frontal (Galynker et
al., 2007)
2.1.2.3. Alcohol
El alcohol (etanol) ejerce sus efectos psicoactivos en el sistema nervioso central a
través de su interacción con múltiples sistemas de neurotransmisores, incluyendo
receptores de GABA, serotonina, opioides y N-Metil-D-Aspartato (NMDA) (un subtipo de
receptor del glutamato). Adicionalmente, el alcohol incrementa los niveles de dopamina en
el estriado a través de un mecanismo indirecto de activación de los receptores GABA, o
inhibición de los receptores NMDA (Camí y Farre, 2003).

P á g i n a | 1 5
Los deterioros vinculados al consumo de alcohol han sido estudiados durante
décadas. Las investigaciones iniciales se centraron en el estudio de los deterioros
asociados con condiciones extremas de alcoholismo como el síndrome de Wernicke-
Korsakoff. Posteriormente, diversos estudios han contribuido a delimitar las alteraciones
neuropsicológicas y de imagen cerebral asociadas al consumo de alcohol en pacientes no
amnésicos.
Alteraciones estructurales
Múltiples estudios de neuroimagen estructural han documentado la existencia de
importantes alteraciones morfológicas en el cerebro de individuos consumidores de
alcohol. Se ha demostrado que estas alteraciones afectan de modo generalizado a diversos
aspectos de la sustancia gris y la sustancia blanca, produciendo atrofia cortical y
reducciones globales del volumen cerebral. Aunque las alteraciones son especialmente
pronunciadas en los lóbulos frontales, el sistema límbico y el cerebelo (Bühler & Mann,
2011; Chanraud et al., 2007; Makris et al., 2008a; Mechtcheriakow et al., 2007; Jang et al.,
2007), otras regiones como la corteza parietal y temporal también han sido encontradas
afectadas (Fein, Shimotsu, Chu, & Barakos, 2009; Gazdzinski et al., 2005).
Alteraciones funcionales
Con la excepción de un estudio (Eckardt, Rohrbaugh, Rio, & Martin, 1990), todos
los estudios llevados a cabo en pacientes dependientes de alcohol en situación de
abstinencia han demostrado reducciones significativas del metabolismo cerebral
fundamentalmente en estructuras frontales y prefrontales (Adams et al., 1993; Dao-
Castellana et al., 1998; Gilman et al., 1990). En un estudio en el que se combinaron técnicas
de PET y resonancia magnética con el objetivo de mejorar la precisión anatómica de las
regiones de interés, Dao-Castellana y colaboradores (1998) detectaron alteraciones del
metabolismo regional en el córtex frontal y prefrontal dorsolateral de consumidores de
alcohol.

P á g i n a | 1 6
2.1.2.4. Éxtasis (MDMA)
El éxtasis, cuyo componente principal es usualmente la 3-
4metilendioximetanfetamina (MDMA), ejerce sus efectos sobre el sistema nervioso central
a través de la inhibición de la recaptación de la serotonina.
Aunque se ha demostrado que la administración de MDMA produce efectos
neurotóxicos selectivos sobre la serotonina en animales (Baumann, Wang, & Rothman,
2007; Green, Mechan, Elliott, O’Shea, & Colado, 2003), los efectos de esta sustancia sobre el
sistema nervioso en humanos han sido ampliamente discutidos (p.e. Green, King, Shortall,
& Fone, 2011). En los últimos años, se ha demostrado la presencia de alteraciones tanto a
nivel estructural como a nivel funcional en consumidores de esta sustancia con o sin
consumo de otras sustancias. Alteraciones consistentes con reducciones significativas de
la disponibilidad de transportadores serotonérgicos en diversas regiones frontales,
temporales, mediales y basales (Parrott, 2012).
Alteraciones estructurales
El consumo crónico de éxtasis ha sido asociado con alteraciones morfológicas
incluso tras importantes períodos de abstinencia en regiones frontales, temporales,
occipitales, cíngulo anterior, cerebelo y ganglios basales. En un estudio llevado a cabo por
Cowan y colaboradores (2003) en el que se comparaban policonsumidores de drogas con
y sin co-abuso de MDMA, aquellos sujetos que consumían MDMA mostraron reducciones
significativas de la sustancia gris cerebral en áreas de la corteza frontal, temporal y
occipital, la corteza cingulada anterior, el tronco cerebral y el cerebelo. Asimismo, en un
reciente estudio en el que se comparó el volumen de materia gris cerebral entre
consumidores con alta y baja exposición de MDMA y anfetaminas y 16 controles sanos, se
encontró que, comparados con los consumidores con bajos niveles de exposición y los
controles sanos, aquellos sujetos que presentaban altos niveles de consumo de estas

P á g i n a | 1 7
sustancias mostraban reducciones significativas de materia gris cerebral en la corteza
orbitofrontal y la corteza anterior del cíngulo.
Alteraciones funcionales
Los estudios del metabolismo cerebral llevados a cabo con PET en consumidores
de éxtasis sugieren una reducción del metabolismo en el giro frontal inferior, los ganglios
basales, el hipocampo, la amígdala y el estriado (Buchert et al., 2001; Obrocki et al., 1999,
2002). Análisis posteriores llevados a cabo por estos autores encontraron asociaciones
significativas entre la duración de la abstinencia y el metabolismo del cíngulo y la amígdala
así como mayores reducciones en aquellos consumidores que habían empezado a utilizar
la sustancia antes de los 18 años de edad, indicando un efecto específico de vulnerabilidad
a la neurotoxicidad de la sustancia en edades tempranas. Sin embargo, no se encontraron
asociaciones significativas entre la cantidad de droga consumida y el metabolismo
cerebral (Buchert et al., 2001; Obrocki et al., 2002).
2.1.2.5. Cannabis
El cannabis, compuesto en su mayor parte por delta-9-tetrahidrocannabinol (THC)
produce sus efectos psicoactivos en el cerebro a través de su acción sobre receptores
cannabinoides CB1. El cannabis, al igual que la heroína, también estimula la producción de
dopamina de manera indirecta a través de la acción de los receptores CB1 sobre las
neuronas de los neurotransmisores de glutamato y GABA en el área tegmental ventral y el
estriatum (Camí & Farré, 2003).
Los estudios llevados a cabo con animales demuestran que la administración de
THC tiene efectos neurotóxicos sobre regiones cerebrales ricas en receptores
canabinoides como el hipocampo, la amígdala o el cerebelo (Pertwee & Ross, 2002). Sin
embargo, en contraste con la literatura animal, los estudios llevados a cabo en humanos
han reportado resultados contradictorios (Martín-Santos et al., 2010).

P á g i n a | 1 8
Alteraciones estructurales
En general no se han encontrado diferencias significativas en el volumen total de
materia gris cerebral entre consumidores de esta sustancia y controles sanos. Sin
embargo, estudios de ciertas regiones cerebrales han encontrado reducciones
significativas de materia gris en el hipocampo, la amígdala y regiones adyacentes en
consumidores abstinentes de esta sustancia (Lorenzetti, Lubman, Whittle, Solowij, &
Yücel, 2010). En general, parece que estas alteraciones se producen en consumidores
severos de esta sustancia (Matochik, Eldreth, Cadet, & Bolla, 2005; Yücel et al., 2008) o en
aquellos que inician su consumo a edades más tempranas (Wilson et al., 2000).
Alteraciones funcionales
Las alteraciones asociadas a los efectos residuales del cannabis consisten en
reducciones significativas del metabolismo cerebral localizadas preferentemente en
regiones frontales y el cerebelo (Sevy et al., 2008; Volkow et al., 1996). En un reciente
estudio llevado a cabo por Sevy y colaboradores (2008) en el que se comparó a un grupo
de 6 consumidores de cannabis en abstinencia con un grupo de 6 controles sanos, los
consumidores de cannabis presentaron reducción del metabolismo cerebral en la corteza
orbitofrontal, el putamen y el precuneus. En este estudio, sólo el metabolismo del córtex
parietal mostró una correlación negativa con la cantidad de uso de esta sustancia.
2.2. Impulsividad y adicciones
La impulsividad es un rasgo estable de la personalidad que varía normativamente
a través de la población normal (Barratt, 1959; Patton, Stanford, & Barratt, 1995) y que ha
sido definida desde la psicología como aquella conducta llevada a cabo con poca o
inadecuada planificación (Evenden, 1999). Este rasgo multifacético de la personalidad, que
en ocasiones puede ser adaptativo (Dickman, 1990), es considerado un rasgo disfuncional

P á g i n a | 1 9
asociado con actos violentos, peligrosos o inadecuados para las demandas de la situación
(Verdejo-García et al., 2008).
Desde de la neuropsicología la impulsividad es equiparada con la "desinhibición",
esto es, la alteración de los mecanismos de control “de arriba a abajo” encargados de
suprimir respuestas automatizadas o impulsadas por la obtención de refuerzos
inmediatos no apropiadas para la situación (Aron, 2007). Estos “mecanismos de control”
pueden verse afectados tanto por lesiones cerebrales como por trastornos mentales dando
lugar a conductas impulsivas. Definida de esta manera, la impulsividad juega un papel
fundamental en los trastornos adictivos. Y es que, las características de personalidad de
una persona podrían influir en las primeras etapas del consumo de la droga por ejemplo
determinando si la persona probará o no la droga o que cantidad tomará y a medida que
avanza el consumo, determinar que la persona persista en su conducta a pesar de las
consecuencias negativas que esta tiene para sí mismo y para los demás (y a pesar de sus
intentos por no hacerlo) o incluso que recaiga tras importantes periodos de abstinencia.
Dentro de la neuropsicología, la impulsividad o desinhibición ha sido evaluada a
través de dos tipos de instrumentos: los cuestionarios de impulsividad y las pruebas
neuropsicológicas que tratan de evaluar la conducta impulsiva o el control inhibitorio.
A continuación se presenta una breve revisión de las medidas que se han utilizado
con más frecuencia en el campo de las adicciones para evaluar la conducta impulsiva y los
principales hallazgos encontrados resultado del uso de las mismas en consumidores en
situación de abstinencia de cocaína, heroína, alcohol, MDMA y cannabis.
2.2.1 Instrumentos de evaluación de la impulsividad
2.2.1.1. Inventarios de personalidad impulsiva
Existe una amplia gama de cuestionarios destinados a medir la impulsividad y
ciertas conductas impulsivas como la búsqueda de sensaciones o la toma de riesgos. En

P á g i n a | 2 0
español contamos con la Escala de Impulsividad de Barratt (Barrat Impulsivity Scale -BIS)
(Oquendo et al., 2001; Patton et al., 1995), la UPPS (Verdejo-García, Lozano, Moya, Alcázar,
& Pérez-García, 2010; Whiteside & Lynam, 2001, 2003), la Escala de Búsqueda de
Sensaciones (Sensation Seeking Scale -SSS) del Cuestionario de Personalidad Zuckerman-
Kuhlman (Zuckerman et al., 1993), adaptada al castellano por Pérez y Torrubia (1986) o la
Escala de Sensibilidad al Castigo y Sensibilidad a la Recompensa (SCSR) (Torrubia, Ávila,
Moltó, & Caseras, 2001). El uso de estos cuestionarios ha mostrado que los consumidores
de diversas sustancias presentan mayores puntuaciones que los controles no
consumidores en diversos índices de impulsividad (ver revisión en Verdejo-García et al.,
2008).
2.2.1.2. Medidas de control inhibitorio
Este componente ha sido definido como la capacidad para inhibir o demorar
respuestas automatizadas, impulsivas o guiadas por el reforzamiento inmediato (Verdejo-
García et al., 2008). Para la evaluación de la impulsividad suelen emplearse tres tipos de
pruebas, aquellas que evalúan la inhibición de respuesta o la capacidad del individuo para
suprimir una respuesta automatizada, medidas de delay-discounting o “descuento
asociado a la demora” y aquellas que tratan de evaluar la impulsividad cognitiva en
términos de toma de decisiones desadaptativas. Dentro de las que permiten evaluar la
inhibición de respuesta o inhibición atencional destacan el test Stroop (Golden, 1978; TEA,
1993), el test de los Cinco Dígitos (Sedó, 2005), las tareas Go – No Go y el Stop Signal Task
(Logan, Cowan, & Davis, 1984). El test de Stroop evalúa la capacidad del individuo para
controlar una respuesta automatizada (leer una palabra) y dar la respuesta adecuada
(decir el color en el que está escrita la palabra) y ha mostrado consistentemente su
capacidad para detectar problemas de atención selectiva e inhibición en consumidores de
distintos tipos de drogas (Verdejo-García, Benbrook, Funderburk, David, Cadet, & Bolla,
2007a; Woicik et al., 2009). El test de los Cinco Dígitos ha demostrado ser una herramienta

P á g i n a | 2 1
útil en la discriminación de los perfiles neuropsicológicos de consumidores de cocaína vs.
opiáceos (Verdejo-García, Perales, & Pérez-García, 2007b) y por último, las tareas Go - No
Go y Stop-Signal han demostrado ser sensibles a la detección de alteraciones en este
componente en consumidores de distintos tipos de drogas (Fillmore & Rush, 2002;
Lawrence, Luty, Bogdan, Sahakian, & Clark, 2009; Monterosso, Aron, Cordova, Xu, &
London, 2005). Para evaluar el “descuento asociado a la demora” o la tendencia de un
individuo a reducir el valor de una recompensa en función del tiempo de espera (Bickel &
Marsch, 2001; Reynolds, 2006), se han utilizado cuestionarios en los que el individuo
selecciona sus preferencias a nivel hipotético (Kirby & Petry, 2004) o tareas en las que el
sujeto tiene que elegir entre pequeñas cantidades entregadas de inmediato o cantidades
mayores entregadas a medio y largo plazo (Reynolds & Schiffbauer, 2004). Por último,
para evaluar el continuo reflexividad-impulsividad se ha recurrido al test de
emparejamiento de figuras conocidas (MFFT-20) (Cairns & Cammock, 2002), la tarea de
recolección de información (Information Sampling Task) (Clark, Robbins, Ersche, &
Sahakian, 2006) y a pruebas de toma de decisiones como la Iowa Gambling Task (Bechara,
Damasio, Damasio, & Anderson, 1994), la Cambridge Gamble Task o la Risky Gains Task
(Rogers et al., 1999a; Rogers et al., 1999b).
2.2.2. Hallazgos relacionados con la impulsividad en consumidores de distintas drogas
2.2.2.1. Cocaína
Inventarios de personalidad impulsiva
El uso de cuestionarios de impulsividad en consumidores de cocaína en situación
de abstinencia ha mostrado que estos pacientes presentan altas puntuaciones de
impulsividad. Las investigaciones llevadas a cabo con estos cuestionarios ha demostrado
que al compararlos con individuos no consumidores, los individuos consumidores de
cocaína presentan altas puntuaciones de urgencia positiva y urgencia negativa (o
tendencia a actuar de forma impulsiva bajo condiciones de estado de ánimo positivo y

P á g i n a | 2 2
negativo respectivamente) (Albein-Urios, Martinez-González, Lozano, Clark, & Verdejo-
García, 2012), falta de premeditación (Fernández-Serrano, Perales, Moreno-López, Pérez-
García, & Verdejo-García, 2012) y búsqueda de sensaciones (Ersche, Turton, Pradhan,
Bullmore, & Robbins, 2010a). Asimismo, el uso de estos cuestionarios en consumidores de
cocaína se ha mostrado como una herramienta de enorme validez en la predicción de la
severidad del consumo, las alteraciones asociadas con el mismo o la predicción de recaídas
(Moeller et al., 2001; Verdejo-García, Bechara, Recknor, & Pérez-García, 2007c)
Medidas de control inhibitorio
Los estudios llevados a cabo en consumidores de cocaína han encontrado
alteraciones en las tareas de inhibición de conducta, descuento asociado a la demora e
impulsividad cognitiva. En este sentido, se ha encontrado que los consumidores de cocaína
en situación de abstinencia presentan más errores durante tareas Go – No Go (Fernández-
Serrano et al., 2012; Verdejo-García et al., 2007b), un mayor tiempo de reacción durante
los ensayos de la tarea Stop Signal (Li, Milivojevic, Kemp, Hong, & Sinha, 2006), mayores
puntuaciones de interferencia durante la realización de pruebas de Stroop (Albein-Urios et
al., 2012; Bolla, Funderburk, & Cadet, 2000) y alteraciones en tareas de delay discounting
(Coffey, Gudleski, Saladin, & Brady, 2003; Heil, Johnson, Higgins, & Bickel, 2006; Kirby &
Petry, 2004; Moeller et al., 2002) y toma de decisiones como el Risky Game o la Iowa
Gambling Task (Stout, Busemeyer, Lin, Grant, & Bonson, 2004; Verdejo-García et al.,
2007b).
De entre las diferentes versiones de la tarea Stroop utilizadas con esta población,
cobran especial relevancia aquellas que han utilizado estímulos emocionales asociados
con la droga (Goldstein et al., 2007). Utilizando estas tareas, diversos estudios han
demostrado que los consumidores de drogas suelen presentar dificultades para filtrar la
información irrelevante o inadecuada cuando ésta tiene que ver con estímulos asociados
con la droga (Nestor, Ghahremani, Monterosso, & London, 2011). Asimismo, se ha descrito

P á g i n a | 2 3
la hipoactivación de los circuitos asociados con el control atencional durante la realización
de esta tarea en estudios de resonancia magnética funcional (Barrós-Loscertales et al.,
2011a; Goldstein et al., 2007).
2.2.2.2. Heroína
Inventarios de personalidad impulsiva
El uso de diversos cuestionarios de impulsividad en consumidores de heroína y
otros opiáceos ha demostrado que estos pacientes presentan elevados niveles de
impulsividad y búsqueda de sensaciones (Cohen et al., 2005; Madden, Petry, Badger, &
Bickel, 1997).
Medidas de control inhibitorio
Se ha encontrado que los consumidores de heroína en situación de abstinencia
presentan altas puntuaciones de interferencia en pruebas de interferencia atencional
(Fernández-Serrano, Pérez-García, Perales, & Verdejo-García, 2010a; Verdejo, Toribio,
Orozco, Puente, & Pérez-García, 2005a), importantes descuentos asociados a la demora
tanto en tareas en las que la recompensa es hipotética como en aquellas en las que es real
(Kirby, Petry, & Bickel, 1999; Kirby & Petry, 2004) y mayor número de errores en tareas
de impulsividad cognitiva (Lee and Pau, 2002) y en tareas de toma de decisiones como la
Iowa Gambling Task (Mintzer, Copersino, & Stitzer, 2005; Verdejo-García et al., 2007b) o la
Cambridge Gamble Task (Fishbein et al., 2007).
2.2.2.3. Alcohol
Inventarios de personalidad impulsiva
Se ha encontrado que los alcohólicos o pacientes con consumo principal de alcohol
en situación de abstinencia muestran índices altos de impulsividad (Mitchell, Fields,
D’Esposito, & Boettiger, 2005), búsqueda de sensaciones (Bjork, Hommer, Grant, &

P á g i n a | 2 4
Danube, 2004) y urgencia negativa (Whiteside & Lynam, 2003). Asimismo, el uso de
cuestionarios de personalidad ha mostrado su capacidad predictiva tanto en la frecuencia
con la que se consume alcohol como en la cantidad consumida o las consecuencias
negativas de dicho consumo (Cyders, Flory, Rainer, & Smith, 2009).
Medidas de control inhibitorio
Las investigaciones llevadas a cabo en esta población han encontrado que los
consumidores de alcohol en situación de abstinencia al ser comparados con individuos no
consumidores cometen más errores en paradigmas Go – No Go (Bjork et al., 2004), tienen
un mayor tiempo de reacción durante tareas de Stop Signal (Goudriaan, Oosterlaan, de
Beurs, & van den Brink, 2006; Lawrence et al., 2009) y presentan mayores índices de
“descuento asociado a la demora” (Mitchell et al., 2005; Petry, 2001). Estos individuos
también presentan alteraciones en tareas de reflexión-impulsividad (Weijers, Wiesbeck, &
Böning, 2001) y toma de decisiones (Dom, De Wilde, Hulstijn, van den Brink, & Sabbe,
2006; Fein, Klein, & Finn, 2004; Fernández-Serrano, Pérez-García, Schmidt Río-Valle, &
Verdejo-García, 2010b).
2.2.2.4. Éxtasis (MDMA)
Inventarios de personalidad impulsiva
El uso de cuestionarios de impulsividad ha permitido encontrar altas puntuaciones
de impulsividad y búsqueda de sensaciones incluso tras 12 meses de abstinencia (Gerra et
al., 2000; Hanson, Luciana, & Sullwold, 2008; Parrott, Sisk, & Turner, 2000).
Medidas de control inhibitorio
Los estudios llevados a cabo en consumidores de éxtasis en situación de
abstinencia han mostrado también la presencia de alteraciones neuropsicológicas durante
la realización de diferentes pruebas de impulsividad (Morgan, Impallomeni, Pirona, &

P á g i n a | 2 5
Rogers, 2006; Quednow et al., 2007). En un estudio correlacional que exploró la asociación
entre la cantidad de MDMA consumida y el rendimiento de un grupo de consumidores
“puros” de MDMA en una amplia batería de pruebas neuropsicológicas, Halpern y
colaboradores (2004) demostraron que aquellos individuos con mayor consumo de esta
sustancia presentaban también mayores puntuaciones de interferencia.
2.2.2.5. Cannabis
Inventarios de personalidad impulsiva
La mayoría de los estudios que han evaluado la personalidad impulsiva en estas
poblaciones han tratado de estudiar los efectos predictivos de estas características sobre
el consumo de la sustancia. Un reciente estudio llevado a cabo por Prince van Leeuwen y
colaboradores (2011) demostró que el uso de la escala de impulsividad de Barratt
permitía predecir el uso y la frecuencia del consumo de esta sustancia. Específicamente, se
encontró que una puntuación alta en la escala de activación conductual se asociaba con
una mayor probabilidad de iniciar el consumo y una puntuación baja en la escala de
inhibición conductual con una mayor probabilidad de persistir en dicho consumo.
Medidas de control inhibitorio
La presencia de alteraciones neuropsicológicas a largo plazo como consecuencia
del consumo de cannabis es todavía hoy objeto de debate. Sin embargo, recientemente se
han descrito alteraciones en consumidores abstinentes de esta sustancia en tareas de
toma de decisiones e impulsividad (Bolla, Eldreth, Matochik, & Cadet, 2005; Pope et al.,
2003; Verdejo-Garcia et al., 2007a).
2.3. Funciones ejecutivas y adicciones
Desde una perspectiva neuropsicológica se considera que la adicción es el
resultado de un conjunto de alteraciones cerebrales que afectan a múltiples sistemas

P á g i n a | 2 6
neurobiológicos y que resultan en disfunciones en procesos motivacionales, emocionales,
cognitivos y conductuales. En los últimos años, la evaluación neuropsicológica de los
individuos drogodependientes ha centrado su atención en las llamadas funciones
ejecutivas, un conjunto integrado de habilidades implicadas en la producción, supervisión
y control de conductas dirigidas a objetivos (Stuss & Knight, 2002; Roberts, Robbins, &
Weiskrantz, 1998) y en la regulación de los estados emocionales que se consideran
adaptativos para la consecución de esos objetivos (Bechara, Damasio, & Damasio, 2000;
Davidson, 2002; Stuss & Alexander, 2000). En el contexto de las adicciones, el principal
objetivo de la neuropsicología es contribuir a dilucidar la naturaleza y extensión de las
alteraciones asociadas con el consumo de drogas mediante el uso de pruebas y tareas
conductuales que impliquen el funcionamiento selectivo de los distintos sistemas
cerebrales implicados en la adicción.
A continuación, se presentarán las principales pruebas utilizadas en la evaluación
neuropsicológica de individuos drogodependientes junto con los principales hallazgos
derivados de su utilización en consumidores de drogas en situación de abstinencia. Como
en apartados anteriores, nos centraremos en el resultado del consumo crónico de cocaína,
heroína, alcohol, MDMA y cannabis, las drogas más consumidas por los participantes de
nuestros estudios.
2.3.1. Evaluación neuropsicológica de las funciones ejecutivas
La aplicación de análisis factoriales y ecuaciones estructurales ha permitido
determinar la existencia de al menos tres componentes diferenciados aunque no
totalmente independientes dentro de las funciones ejecutivas (Fisk & Sharp, 2004; Miyake
et al., 2000). Estos componentes han sido definidos como actualización, inhibición de
respuestas y cambio. Además, recientemente se ha propuesto que la toma de decisiones
constituiría un cuarto componente independiente dentro de las funciones ejecutivas. Una
propuesta avalada por la evidencia que demuestra la existencia de importantes

P á g i n a | 2 7
alteraciones en los procesos de toma de decisiones en individuos drogodependientes y el
hecho de que estas no correlacionen con la ejecución de estos individuos en tareas que
implicadas componentes de actualización, inhibición y cambio (Bechara et al., 2001; Grant,
Contoreggi, & London, 2000; Verdejo-García & Pérez-García, 2007). Asimismo, en los
últimos años, los modelos neuropsicológicos de la adicción han enfatizado la importancia
de los factores emocionales en los procesos adictivos (Goldstein & Volkow, 2002; Redish,
Jensen, & Johnson, 2008; Verdejo-García & Bechara, 2009).
2.3.1.1. Actualización
El componente de actualización implica la monitorización, actualización y
manipulación de la información “on line” en la memoria operativa (Miyake et al., 2000).
Recientemente se ha propuesto que este componente incluye los procesos de memoria de
trabajo, fluidez y razonamiento (Verdejo-García & Pérez-García, 2007).
La memoria de trabajo es un sistema que permite el almacenamiento,
manipulación y actualización temporal de la información en el cerebro (M D’Esposito et al.,
1995). Tradicionalmente se ha evaluado utilizando algunos subtests de la escala WAIS
(Wechsler Adult Intelligence Scale, Wechsler, 1997) y otros instrumentos como la prueba
de Span visual de la Escala de Memoria Wechsler o las tareas n-back (Watter, Geffen, &
Geffen, 2001). Estas tareas han demostrado ser eficaces en la detección de alteraciones en
procesos de memoria de trabajo en consumidores de diversas drogas tanto en estudios
conductuales como en estudios de neuroimagen funcional (Bustamante, Barrós-
Loscertales, Ventura-Campos, Sanjuán, Llopis, Parcet, & Avila, 2011a; Tomasi, Goldstein,
Telang, Maloney, Alia-Klein, Caparelli, & Volkow, 2007). Podemos definir la fluidez como la
capacidad del individuo para iniciar su conducta de forma espontánea y creativa en
respuesta a una orden novedosa. Para evaluar este proceso se han empleado el Test de
fluidez verbal (FAS) (Lezak, 2004), Animales, Frutas y Herramientas y el Ruff Figural
Fluency Test (RFFT) (Ruff, 1996). Por último, el razonamiento analógico consiste en

P á g i n a | 2 8
obtener una conclusión a partir de premisas sobre las que se establece una comparación o
analogía entre elementos o conjuntos de elementos distintos. Para la evaluación de este
dominio tradicionalmente se ha recurrido al subtest de Semejanzas del WAIS.
2.3.1.2. Control inhibitorio
La evaluación del control inhibitorio ha sido ampliamente descrita en el apartado
de evaluación de la impulsividad en individuos drogodependientes. Brevemente, este
componente ha sido evaluado a través de tareas de inhibición de respuesta, “descuento
asociado a la demora” y tareas de toma de decisiones.
2.3.1.3. Flexibilidad cognitiva
La flexibilidad cognitiva es la capacidad del individuo para reestructurar su
conocimiento y adaptar su respuesta a las exigencias cambiantes del ambiente (Spiro &
Jehng, 1990). Se trata también de un componente multidimensional que ha sido estudiado
a través de distintos índices incluyendo pruebas que miden la respuesta del individuo ante
el cambio de las reglas de una tarea, el criterio de respuesta o el set atencional y tareas de
reversal learning que miden la capacidad del individuo para cambiar su respuesta en
función de cambios en los patrones de reforzamiento.
Para la evaluación de la respuesta del individuo ante el cambio de set se ha
recurrido a tareas como la Prueba de Categorías (DeFilippis, 2002) o el Test de
Clasificación de Tarjetas de Wisconsin (WCST) (Grant & Berg, 1948). La prueba de
Categorías es una tarea informatizada que presenta tamaños del efecto considerables en la
discriminación del rendimiento de consumidores y controles (Verdejo-García & Pérez-
García, 2007) y ha demostrado buenos valores de predicción del resultado del tratamiento
en consumidores de cocaína (Turner et al., 2009).
2.3.1.4. Toma de decisiones

P á g i n a | 2 9
La toma de decisiones es la habilidad para seleccionar de entre un conjunto de
posibles alternativas aquella que resulte más beneficiosa para el individuo. Una de las
pruebas más utilizadas dentro del campo de la neuropsicología es la Iowa Gambling Task
(Bechara et al., 1994). Esta prueba ha mostrado ser sensible a la detección de alteraciones
en la toma de decisiones en consumidores de diversas drogas. Otras tareas empleadas en
la evaluación de la toma de decisiones son el Juego de Datos (Game of Dice Task) o la
Cambridge Gamble Task. Estas tareas pueden ser descritas como pruebas más ecológicas
ya que el individuo tiene información sobre las potenciales consecuencias de sus acciones
(Brand, Labudda, & Markowitsch, 2006). En los últimos años, diversos estudios han
mostrado que la utilización de índices de toma de decisiones en condiciones ambiguas y de
riesgo son de gran utilidad en la predicción de recaídas (Bowden-Jones, McPhillips,
Rogers, Hutton, & Joyce, 2005; Passetti, Clark, Mehta, Joyce, & King, 2008; Paulus, Tapert,
& Schuckit, 2005).
2.3.1.5. Procesos emocionales
Las investigaciones llevadas a cabo en pacientes drogodependientes han
demostrado que estos pacientes presentan alteraciones tanto en la percepción como en la
experiencia emocional.
Uno de los aspectos claves del funcionamiento emocional es la capacidad para
identificar y reconocer señales emocionales en las caras de otras personas. Se ha
demostrado que el reconocimiento emocional es fundamental para la conducta prosocial,
la socialización y la interacción normal (Blair, 2003) y que la alteración de dicho proceso
cursa con alteraciones en la experiencia emocional (Calder & Young, 2005). El paradigma
más utilizado en la evaluación de la percepción emocional consiste en la presentación de
un conjunto de fotografías de personas que expresan diferentes emociones que la persona
tiene que identificar. De entre las diferentes tareas, la prueba más utilizada dentro del
campo de las adicciones ha sido el test de Ekman. Esta prueba evalúa mediante la

P á g i n a | 3 0
presentación de un conjunto de estímulos procedentes del banco de “Expresiones Faciales
Emocionales: Estímulos y Tests” (Facial Expressions of Emotion: Stimuli and Tests –
FEEST) (Young, Perrett, Calder, Sprengelmeyer & Ekman, 2002) la capacidad del individuo
para identificar expresiones faciales representativas de las seis emociones básicas
(felicidad, tristeza, miedo, asco, ira y sorpresa).
2.3.2. Hallazgos neuropsicológicos relacionados con las funciones ejecutivas en
consumidores de distintas drogas
Una revisión de los estudios llevados a cabo hasta el momento en consumidores de
drogas de diferente tipo, indica que aunque existen ciertos mecanismos neuropsicológicos
que parecen verse afectados por todas las drogas de abuso, también existen ciertas
alteraciones que podrían ser especificas al consumo de cada sustancia (Fernández-
Serrano, Pérez-García, & Verdejo-García, 2011). En este sentido, aunque todas las drogas
han sido asociadas con la presencia de alteraciones en los procesos de memoria,
actualización, toma de decisiones y procesamiento emocional, el uso de psicoestimulantes
y alcohol ha sido asociado con el control inhibitorio y la flexibilidad, el consumo de alcohol
y MDMA con el procesamiento espacial, la velocidad perceptiva y la atención selectiva, el
uso de cannabis y metanfetaminas con la presencia de déficits en memoria prospectiva y
por último, el uso de cannabis y MDMA ha sido asociado con la presencia de alteraciones
en velocidad de procesamiento y planificación.
Por otra parte, la mayoría de las investigaciones que han tratado de evaluar la
percepción emocional en sujetos consumidores de drogas se han centrado en los efectos
producidos por el consumo de alcohol, y en la mayor parte de los casos en el estudio de la
habilidad para estimar la intensidad de las emociones expuestas pero no la exactitud o
precisión en el reconocimiento emocional. De entre aquellos estudios dirigidos a estudiar
la eficacia del reconocimiento emocional en estas poblaciones, aunque algunos estudios no
han encontrado diferencias significativas entre consumidores de drogas y controles

P á g i n a | 3 1
(Salloum et al., 2007; Woicik et al., 2009), la mayoría de los estudios llevados han
encontrado diferencias significativas en el reconocimiento de las emociones en general y
de las emociones negativas en particular. Asimismo, los estudios que han tratado de
evaluar la experiencia emocional en pacientes drogodependientes coinciden en señalar
que estos pacientes se activan menos y muestran mayor control que los individuos no
consumidores (Aguilar de Arcos et al., 2008; Aguilar de Arcos, Verdejo-García, Peralta-
Ramírez, Sánchez-Barrera, & Pérez-García, 2005), resultados que coinciden con la
creciente evidencia que demuestra que los individuos drogodependientes presentan una
sobreactivación de las regiones cerebrales implicadas en el procesamiento emocional de
recompensas en respuesta a estímulos asociados con la sustancia de consumo (Childress
et al., 1999; George et al., 2001; Kilts, Gross, Ely, & Drexler, 2004; Tapert, Brown, Baratta, &
Brown, 2004) y una reducción de la actividad cerebral en estas mismas regiones en
respuesta a reforzadores naturales como imágenes de contenido sexual (Asensio et al.,
2010; Garavan et al., 2000).
2.3.2.1. Cocaína
El consumo de cocaína ha sido consistentemente asociado con la presencia de
alteraciones neuropsicológicas en los procesos cognitivos de atención y memoria,
funcionamiento ejecutivo y procesos emocionales. Una revisión de los estudios llevados a
cabo en los últimos años indica que las alteraciones más frecuentemente encontradas en
consumidores de cocaína y policonsumidores con consumo principal de cocaína son las
alteraciones de los procesos de memoria, flexibilidad e inhibición (Fernández-Serrano et
al., 2011; Ruth Janke van Holst & Schilt, 2011). La alteración de la memoria de trabajo
constituye una de las características más frecuentemente reportadas en estudios de
cocaína (Fernández-Serrano et al., 2011) y ha sido recientemente asociada con la
neurotoxicidad de esta sustancia (Albein-Urios et al., 2012). Asimismo se han descrito
alteraciones en atención (Pace-Schott et al., 2008; Verdejo-García & Pérez-García, 2007;

P á g i n a | 3 2
Woicik et al., 2009), memoria y aprendizaje verbal (De Oliveira et al., 2009; Fox, Jackson, &
Sinha, 2009; Goldstein et al., 2004; Jovanovski, Erb, & Zakzanis, 2005; Verdejo-García &
Pérez-García, 2007; Woicik et al., 2009), fluidez (Verdejo-García & Pérez-García, 2007),
inhibición (Verdejo-García & Pérez-García, 2007), flexibilidad cognitiva (Goldstein et al.,
2004; Woicik et al., 2009), toma de decisiones (Fernández-Serrano et al., 2010b; Verdejo-
Garcia et al., 2007a) y reconocimiento emocional (Fernández-Serrano, Lozano, Pérez-
García, & Verdejo-García, 2010c; Verdejo-García, Rivas-Pérez, Vilar-López, & Pérez-García,
2007d).
2.3.2.2. Heroína
La mayoría de los estudios llevados a cabo en consumidores de opiáceos o heroína
defienden la existencia de alteraciones en los dominios de fluencia, inhibición y toma de
decisiones (van Holst & Schilt, 2011). En una revisión llevada a cabo en 2007, Gruber y
colaboradores concluyeron que el consumo de opiáceos producía alteraciones a largo
plazo en los procesos de atención, concentración, memoria y habilidades visuo-
perceptivas. Otros autores han descrito alteraciones en los dominios de memoria de
trabajo, flexibilidad cognitiva e inhibición (Brand, Rothbauer, Driessen, & Markowitsch,
2008; Prosser et al., 2008). Asimismo se han descrito alteraciones en fluidez (Fernández-
Serrano et al., 2010a), razonamiento (Verdejo et al., 2005a), toma de decisiones (Brand et
al., 2008; Verdejo-García & Pérez-García, 2007) y procesos emocionales (Kornreich et al.,
2003; Wang et al., 2010).
2.3.2.3. Alcohol
El consumo de alcohol ha sido consistentemente asociado con la presencia de
alteraciones neuropsicológicas en los procesos de atención y memoria, funcionamiento
ejecutivo y procesos emocionales. Una reciente revisión de los estudios que han tratado de
evaluar las alteraciones neuropsicológicas producidas por el consumo de alcohol en
individuos drogodependientes en situación de abstinencia concluye que las alteraciones

P á g i n a | 3 3
más frecuentemente reportadas son las encontradas en memoria de trabajo, memoria
verbal, inhibición y habilidades visuo-perceptivas (van Holst & Schilt, 2011). El consumo
de alcohol y psicoestimulantes han sido asociado con la presencia de alteraciones del
control inhibitorio y la flexibilidad cognitiva y el consumo de esta sustancia con MDMA ha
sido asociado con alteraciones en tareas de procesamiento espacial, velocidad perceptiva y
atención selectiva (Fernández-Serrano et al., 2011). En general, aunque parece que se
produce una mejora de los procesos cognitivos tras periodos de abstinencia relativamente
breves (Manning et al., 2008; Moriyama, Muramatsu, Kato, Mimura, & Kashima, 2006;
Tedstone & Coyle, 2004; Zinn, Stein, & Swartzwelder, 2004), los procesos de
funcionamiento ejecutivo se han encontrado alterados incluso tras varios años de
abstinencia (Davies et al., 2005; Fein et al., 2004).
A nivel emocional, se ha encontrado que los alcohólicos tienden a sobreestimar la
intensidad de la emoción que aparece en las expresiones faciales de felicidad, ira y asco
(Foisy et al., 2007; Kornreich et al., 2001; Townshend & Duka, 2003) y que presentan
alteraciones en el reconocimiento de las emociones en general y de las emociones
negativas en particular (Frigerio, Burt, Montagne, Murray, & Perrett, 2002; Townshend &
Duka, 2003).
2.3.2.4. Éxtasis (MDMA)
El consumo de éxtasis o MDMA ha sido asociado con alteraciones en todos los
procesos neuropsicológicos descritos. Parece que las alteraciones más frecuentemente
encontradas son aquellas que se producen en tareas de memoria, concretamente en
aquellas que evalúan memoria verbal (Kalechstein, De La Garza, Mahoney, Fantegrossi, &
Newton, 2007; Parrott, 2012; Ruth Janke van Holst & Schilt, 2011). También se han
descrito alteraciones en el reconocimiento emocional (Yip & Lee, 2006).
En los últimos años, varios estudios han llamado la atención sobre el efecto que el
co-abuso de otras sustancias y en especial del cannabis podría tener sobre los efectos

P á g i n a | 3 4
neuropsicológicos asociados con el consumo de éxtasis (p.e. Schulz, 2011). En este sentido,
mientras que el uso de MDMA y alcohol ha sido asociado con la presencia de alteraciones
en los procesos de atención selectiva, procesamiento espacial y velocidad perceptiva en
consumidores de estas sustancias, el uso de MDMA y cannabis ha sido asociado con la
planificación y la velocidad de procesamiento (Fernández-Serrano et al., 2011).
2.3.2.5. Cannabis
Todavía hoy existe un importante debate acerca de las alteraciones producidas por
el consumo de cannabis. Aunque el consumo de esta sustancia ha sido consistentemente
asociado con la presencia de alteraciones neuropsicológicas a corto plazo sobre todo en
los procesos de memoria y atención, las revisiones publicadas hasta el momento de los
estudios que han tratado de evaluar los efectos a largo plazo de esta sustancia coinciden
en señalar la ausencia de alteraciones significativas en consumidores de esta sustancia en
situación de abstinencia (Grant, Gonzalez, Carey, Natarajan, & Wolfson, 2003; van Holst &
Schilt, 2011). En los últimos años, algunos autores han reportado alteraciones tras
importantes periodos de abstinencia en consumidores severos de la sustancia (Bolla et al.,
2005; Montgomery, Seddon, Fisk, Murphy, & Jansari, 2012; Verdejo-García & Pérez-García,
2007) y en aquellos que iniciaron su consumo en la adolescencia (Fontes et al., 2011;
Medina et al., 2007; Pope et al., 2003). Parece que estas alteraciones se producen
fundamentalmente en los procesos de toma de decisiones, la formación de conceptos y la
planificación (Crean, Crane, & Mason, 2011).

P á g i n a | 3 5
JUSTIFICACIÓN Y OBJETIVOS [ ]

P á g i n a | 3 6
La aplicación de las técnicas de neuroimagen cerebral al ámbito de las
drogodependencias ha permitido en los últimos años examinar “in vivo” los efectos agudos
de la administración de sustancias psicoactivas, los correlatos cerebrales del deseo intenso
por consumir las drogas (craving), o las alteraciones a largo plazo en regiones cerebrales y
sistemas neuroquímicos implicados en el consumo crónico, la dependencia y la recaída.
En los últimos años, un aspecto metodológico que ha suscitado un creciente interés
es la dirección causal de la relación entre las alteraciones neuropsicológicas encontradas y
el consumo de drogas, ya que diversos estudios han puesto de manifiesto que las
alteraciones neuropsicológicas (en especial las alteraciones de las funciones ejecutivas)
pueden predecir el inicio del consumo de drogas y contribuir al desarrollo de las
adicciones (Giancola & Tarter, 1999; Verdejo-García et al., 2008). Sin embargo, esta noción
no es incompatible con la evidencia empírica que indica que el consumo prolongado de
drogas puede generar alteraciones neuropsicológicas en individuos drogodependientes.
Por otro lado, el estudio de las alteraciones neuropsicológicas asociadas al consumo de
drogas y sus correlatos neurobiológicos puede verse afectado por varias limitaciones
metodológicas inherentes al contexto de la investigación en drogodependencias que
dificultan en gran medida la interpretación de los resultados obtenidos y que pueden dar
lugar a inconsistencias y contradicciones. En este sentido, es importante destacar el hecho
de que la mayoría de los individuos drogodependientes son policonsumidores de drogas y
suelen presentar patrones de intensidad, frecuencia y duración del consumo o de
duración de la abstinencia muy heterogéneos. El policonsumo y en particular la
combinación de drogas ilegales con alcohol y, en ocasiones, con medicamentos y
sustancias no reguladas, se ha convertido en la pauta dominante del consumo de drogas en
Europa (EMCDDA, 2011), por lo que la investigación de los efectos diferenciales de
distintas drogas de abuso está frecuentemente limitada por el hecho de que la práctica
totalidad de los individuos drogodependientes no son consumidores exclusivos de una

P á g i n a | 3 7
única sustancia, sino policonsumidores de diversas sustancias (Fernández-Serrano et al.,
2011; Verdejo-García et al., 2004).
Aunque puede resultar complejo afrontar diseños de investigación capaces de
controlar el conjunto de las limitaciones mencionadas, el conocimiento y la consideración
de estas limitaciones en la interpretación de los resultados puede incrementar
significativamente la capacidad explicativa de las investigaciones en el ámbito de las
drogodependencias.
En el contexto de aportar conocimientos sobre esta temática y superar las
limitaciones existentes en estudios previos de adicción, la presente Tesis Doctoral tiene
como Objetivo General el estudio de las alteraciones cerebrales estructurales y
funcionales asociadas con variables de personalidad que incrementan la predisposición al
riesgo de consumo y la dependencia, con el consumo crónico de diferentes drogas y con el
funcionamiento neuropsicológico y emocional de un grupo de pacientes
drogodependientes en situación de abstinencia.
De este Objetivo General se derivaron tres Objetivos Específicos, que se articularon
a través de tres estudios. En primer lugar, se abordaron las posibles alteraciones
estructurales de pacientes con consumo principal de cocaína. En este sentido, el Primer
Objetivo fue estudiar la asociación entre una medida multidimensional de impulsividad
rasgo y las alteraciones de la estructura cerebral presentadas por un grupo de pacientes
policonsumidores de drogas con consumo principal de cocaína. Una vez estudiadas las
alteraciones estructurales encontradas en esta población y su asociación con medidas de
personalidad, los dos siguientes estudios se dirigieron a investigar las alteraciones
funcionales asociadas con la severidad del consumo (estudio 2) y las alteraciones
neuropsicológicas encontradas en esta población (estudio 3). Por tanto, el Segundo
Objetivo fue estudiar la asociación entre la severidad del consumo de diversas drogas y el
metabolismo cerebral en reposo de un grupo de pacientes policonsumidores de drogas en

P á g i n a | 3 8
situación de abstinencia y el Tercer Objetivo estudiar la asociación del funcionamiento
neuropsicológico y el metabolismo cerebral en situación de reposo en este mismo grupo
de policonsumidores de drogas.
Las Hipótesis que se derivaron de cada uno de estos Objetivos fueron:
Para el Primer Objetivo: Esperamos encontrar una asociación significativa entre la
personalidad impulsiva y el volumen cerebral de la corteza prefrontal y los ganglios
basales específica de los individuos consumidores de cocaína.
Para el Segundo Objetivo: Esperamos encontrar una asociación generalizada de la
severidad del consumo de todas las drogas estudiadas con el metabolismo cerebral de
regiones fronto-estriadas y el cerebelo en individuos policonsumidores, así como una
asociación específica del consumo de cocaína con la corteza parietal y del de heroína con la
corteza temporal una vez controlado el co-abuso de las demás drogas consumidas.
Para el Tercer Objetivo: Esperamos encontrar una asociación entre el rendimiento
en pruebas de función ejecutiva “frías” y el metabolismo cerebral de la corteza prefrontal
dorsolateral, la parte anterior del cíngulo y estructuras temporales y entre pruebas de
función ejecutiva “caliente” y el metabolismo cerebral de la corteza orbitofrontal, la parte
anterior del cíngulo y el sistema límbico en el grupo de policonsumidores.
El seguimiento de estos tres Objetivos e Hipótesis ha dado lugar a la realización de
tres trabajos de investigación y a la publicación de tres Manuscritos en revistas
internacionales de alto impacto tras un proceso de revisión por pares y cuyas referencias
se citan a continuación:
Moreno-López, L., Catena, A., Fernández-Serrano, M.J., Delgado-Rico, E., Stamatakis,
E.A., Pérez-García, M., Verdejo-García, A. (2012). Trait impulsivity and prefrontal
gray matter reductions in cocaine dependent individuals. Drug and Alcohol
Dependence, en prensa.

P á g i n a | 3 9
Moreno-López, L., Stamatakis, E.A., Fernández-Serrano, M.J., Gómez-Río, M.,
Rodríguez-Fernández, A., Pérez-García, M., Verdejo-García, A. (2012) Neural
Correlates of the Severity of Cocaine, Heroin, Alcohol, MDMA and Cannabis Use
in Polysubstance Abusers: A Resting-PET Brain Metabolism Study. PLoS ONE,
en prensa.
Moreno-López, L., Stamatakis, E.A., Fernández-Serrano, M.J., Gómez-Río, M.,
Rodríguez-Fernández, A., Pérez-García, M., Verdejo-García, A. (2012) Neural
correlates of hot and cold executive functions in polysubstance addiction:
Association between neuropsychological performance and resting-PET brain
metabolism. Psychiatry Research: Neuroimaging, en prensa.

P á g i n a | 4 0

P á g i n a | 4 1
[MEMORIA DE TRABAJOS]

P á g i n a | 4 2

P á g i n a | 4 3
* ARTÍCULO 1
Trait impulsivity and prefrontal gray matter reductions in cocaine dependent individuals
* Moreno-López, L., Catena, A., Fernández-Serrano, M. J., Delgado-Rico, E., Stamatakis, E. A.,
Pérez-García, M., & Verdejo-García, A. (2012). Trait impulsivity and prefrontal gray matter
reductions in cocaine dependent individuals. Drug and Alcohol Dependence, en prensa.

P á g i n a | 4 4
1. Introduction
Cocaine addiction is a worldwide major public health problem for which current
prevention and treatment options are not fully satisfactory (EMCDDA, 2009; Substance
Abuse and Mental Health Services Administration, 2010). Neuroimaging studies in
cocaine-dependent individuals have revealed significant brain volume reductions, most
notably in a priori selected regions of interest, such as the striatum, the amygdala or the
prefrontal cortex (Barrós-Loscertales et al., 2011; Makris et al., 2004; Matochik et al.,
2003; Tanabe et al., 2009). These regions are thought to contribute to critical aspects of
the addictive cycle, including reinforcement learning, craving, and inhibitory control
(Koob and Volkow, 2010). Nonetheless, the findings from nonbiased automated
techniques, such as Voxel Based Morphometry (VBM), have yielded inconsistent results.
Despite previous positive findings showing significant gray matter (GM) reductions in
cocaine-dependent individuals, including the medial prefrontal cortex, superior temporal
cortex, insula, thalamus and cerebellum (Franklin et al., 2002; Sim et al., 2007), a recent
VBM study failed to detect any structural change (GM or white matter –WM– reductions)
in cocaine users compared to non-drug using controls (Narayana et al., 2010). Perhaps
more critically, most of these studies have failed to find any correlation between estimates
of drug use patterns (e.g., amount, duration or age at onset) and GM and WM reductions
(Franklin et al., 2002; Makris et al., 2004; Matochik et al., 2003; but see Ersche et al., 2011).
This apparent lack of association between cocaine exposure and brain attrition raises the
possibility that certain personality traits, such as impulsivity, may relate to GM and WM
abnormalities in cocaine-dependent individuals.
Impulsivity is viewed as a multifaceted trait that varies normally across the
population, but that in high levels may predispose to a range of dysfunctional behaviors,
including addiction (Cyders et al., 2007; Verdejo-García et al., 2008). Animal studies have
shown that increased impulsivity is associated with reduced dopamine receptors

P á g i n a | 4 5
availability and cocaine use escalation and progression to dependence (Belin et al., 2008;
Dalley et al., 2007). In humans, high levels of trait impulsivity (indexed with the Barratt
Impulsivity Scale) are associated with lower midbrain dopamine autoreceptor binding,
and greater amphetamine-induced dopamine release in the striatum (Buckholtz et al.,
2010). In addition, trait impulsivity is negatively correlated with orbitofrontal cortex
(OFC) GM volumes in healthy individuals (Matsuo et al., 2009). Nonetheless, in addicted
individuals, continued use of stimulants is thought to further exacerbate impulsive traits
(Ersche et al., 2010), and to possibly modify its neural underpinnings. In fact, the
association between trait impulsivity levels and striatal dysfunction is stronger in
methamphetamine-dependent individuals compared to healthy controls (Lee et al., 2009).
Conversely, Ersche et al. (2011) failed to find significant correlations between particular
aspects of trait impulsivity (impulsive reward-seeking) and GM reductions in cocaine-
dependent subjects. Therefore, more studies are needed to specifically explore if, as
expected, different facets of trait impulsivity are specifically linked to GM volumes in
frontostriatal regions in cocaine-dependent individuals.
The aims of this study were (i) to use whole-brain VBM analyses to examine
possible GM and WM reductions in a sample of currently abstinent (>1 month) cocaine
dependent individuals, as compared to a non-drug using control group; and (ii) to
measure differences in the way impulsivity relates to GM and WM volumes in cocaine
users vs. controls. Because impulsive personality in the normal population is negatively
associated with GM volume in the OFC (Matsuo et al., 2009), which is also impacted by
lifetime cocaine use (Alia-Klein et al., 2011), we expected cocaine-dependent individuals to
have reduced GM volumes in the OFC and richly interconnected regions, such as insula,
amygdala, striatum, and WM adjacent to these regions (Bechara, 2005). Because
progression of cocaine use is thought to provoke neuroadaptations from the ventral
striatum to the dorsal striatum, and from the OFC to more extensive prefrontal regions
like the anterior cingulate and dorsolateral prefrontal cortex (Koob and Volkow, 2010), we

P á g i n a | 4 6
expected impulsivity to be uniquely associated with more lateral aspects of the prefrontal
cortex and more posterior aspects of the striatum in cocaine-dependent individuals.
2. Materials and Methods
2.1. Participants
Thirty-eight cocaine-dependent individuals, mean age=29.58, SD=6.53, and 38
non-drug using controls, mean age=31.08, SD=5.14, participated in this study. All
participants were male due to the low prevalence of women entering drug treatment
during the recruitment period. Cocaine users were recruited in an inpatient therapeutic
community (“Proyecto Hombre”), in the city of Granada, Spain. All of them reported
cocaine as their main drug of choice and the one for which they requested treatment.
Clinical interviews based on Diagnostic and Statistical Manual version IV (DSM-IV) criteria
confirmed cocaine dependence diagnosis; nonetheless, they also had regular use of
tobacco, alcohol, cannabis and MDMA (see Table 1). To enter the study, cocaine users had
to be abstinent for at least 30 days (for any drug but tobacco), as confirmed by weekly
urine tests; in this way, we could rule out acute and residual effects of previously used
drugs on brain structure, with the exception of nicotine (84.2% of cocaine-dependent
individuals and 44.7% of controls were current smokers). None of the cocaine patients
were currently following pharmacological substitution treatments. Potential participants
who had previously been diagnosed with any disorder from DSM- IV Axes I and II (other
than substance dependence), or had neurological or systemic diseases affecting central
nervous system (CNS) functioning were excluded.
Non-drug using controls were recruited through adverts distributed by a local
employment agency, and therefore they were also matched to cocaine participants in
terms of unemployment status. Selection criteria for control participants were: (i) absence
of current or past substance use, excluding past or current social drinking (less than ten
standard alcohol units per week), (ii) absence of documented major psychiatric disorders,

P á g i n a | 4 7
(iii) absence of documented head injury or neurological disorder, and (iv) not being taking
medication with effects on the CNS.
For both groups, evidence of stroke or space-occupying lesions observed on
conventional clinical MR images, any contraindications to MRI scanning (including
claustrophobia and implanted ferromagnetic objects), and history of loss of consciousness
(LOC) for longer than 30 minutes or LOC with any neurological consequence were
exclusionary.
2.2. Instruments and assessment procedures
The study was approved by the Ethics Committee for Research in Humans of the
University of Granada. All participants signed an informed consent form certifying their
voluntary participation. Controls, but not patients, received a €40 compensation for
participating in the study. Assessments were conducted across two sessions separated by
less than one week. During the first session we administered the Interview for Research
on Addictive Behavior, the Hamilton Rating Scales for Depression and Anxiety (HAM-D
and HAM-A), and the UPPS-P Impulsive Behavior Scale, along with a battery of cognitive
tests which results will be reported separately. The second session involved the MRI
scanning. The MRI scans lasted approximately 6 minutes.
2.2.1. Patterns of Drug Use
Data regarding lifetime amount and duration of use of the different drugs was self-
reported by participants and collected using the Interview for Research on Addictive
Behavior (Verdejo-García et al., 2005). This interview provides an estimation of monthly
use of each substance during regular use (amount per month) and total duration of use of
each substance (in years). The descriptive scores for these variables in the sample are
presented in Table 1.

P á g i n a | 4 8
Table 1 Descriptive information about patterns of drug use in cocaine-dependent individuals.
Substance Ever used (%) Unit Amount per month Duration of use Abstinence
Cocaine 100 # Grams 18.92 ± 29.46 4.05 ± 3.07 36.76 ± 22.60
Cannabis 76.31 # Joints 109.90 ± 111.40 2.35 ± 3.17 147.45 ± 203.66
MDMA 50 # Tablets 9.29 ± 10.12 2.27 ± 1.90 227.68 ± 225.78
Alcohol 100 # SAU 107.58 ± 114.03 7.38 ± 5.77 33.11 ± 21.77
Tobacco 86.8 # Cigarettes 488.48 ± 307.40 9.19 ± 7 7.8 ± 15.82
Note: Mean ± Standard deviation of the mean. Duration of use is expressed in years. Abstinence duration is expressed in number of weeks.

P á g i n a | 4 9
2.2.2. Trait Impulsivity
We used the Spanish version of the UPPS-P Scale (Verdejo-García et al., 2010;
Whiteside and Lynam, 2001). This is a 59-item self-report inventory designed to measure
five distinct personality pathways to impulsive behavior: sensation seeking, (lack of)
perseverance, (lack of) premeditation, negative urgency, and positive urgency. The first 4
dimensions were included in the original version of the UPPS scale (Whiteside and Lynam,
2001); the fifth dimension has been included based on recent work by (Cyders et al., 2007;
Smith et al., 2007). Sensation seeking (12 items) incorporates two aspects: 1) a tendency
to enjoy and pursue activities that are exciting, and 2) an openness to trying new
experiences that may or may not be dangerous; (lack of) perseverance (10 items) refers to
the individual’s ability to remain focused on a task that may be boring or difficult; (lack of)
premeditation (11 items) refers to the tendency to think and reflect on the consequences
of an act before engaging in that act; and finally urgency (12 items) refers to the tendency
to experience strong impulses under conditions of negative affect (negative urgency, 12
items) or positive affect (positive urgency, 14 items). Each item on the UPPS is rated on a
4-point scale ranging from 1 (strongly agree) to 4 (strongly disagree). We obtained the
total scores of each of these five UPPS-P dimensions for analysis.
2.3. MRI acquisition
Participants were scanned on a 3T whole body MRI scanner (Phillips Achieva)
operating with 8 channels phased-array head coil for reception. For each participant, a T1-
weighted 3D volume was acquired using a T1-weighted 3D-turbo-gradient-echo sequence
(3D-TFE), in sagittal orientation with 0.94x0.94x1.0 mm resolution (160 slides,
FOV=240x240 mm2, matrix 256x256x160) with repetition time of 8 ms, echo time 4 ms,
inversion delay = 1022.6264 ms, flip angle of 8°, band with 191 Hz/pixel. The sequence
was optimal for reducing motion sensitivity, susceptibility artifacts and field
inhomogeneities.

P á g i n a | 5 0
2.4. Image analysis
Before automatic preprocessing, the images were checked for artifacts and
manually aligned to the AC-PC line. Data were processed and analyzed using SPM8
(http://www.fil.ion.ucl.ac.uk/spm) and VBM8 toolbox (http://dbm.neuro.unijena.
de/vbm.html), for which we used the default parameters. Thus, within a unified
segmentation model (Ashburner and Friston, 2005), images were corrected for biasfield
inhomogeneities, registered using linear (12 parameters affine) and non-linear
transformations (warped), and tissue was clustered into GM, WM, and cerebrospinal fluid
(CSF). Segments were further refined by using adaptive maximum a posteriori
estimations, which account for partial volume effects, and by applying a hidden Markov
random field model, as implemented in VBM8. Importantly, to preserve local GM/WM
values, we multiplied the segments by the Jacobian determinants of the deformation field
to create modulated images. Segments were smoothed by an 8 mm full-width at half-
maximum (FWHM) using an isotropic Gaussian kernel. Afterwards, we conducted analysis
on modulated GM and WM segments.
2.4.1. Global effects of patterns of drug use
Total gray and white matter volumes (TGMV, TWMV) were computed by adding up
the voxel values of GM/WM segments, including cerebellum and sub-cortical structures.
Next, the total brain volume (TBV=TGMV+TWMV) was used for computing the TGMV/TBV
ratios, which served to evaluate the global effects of patterns of drug use on brain tissue
volumes using the backward step-wise procedure in the general linear model, including as
predictors age, years of education, cocaine amount per month during regular use, duration
of cocaine use, and age of onset of cocaine use. T-tests were used to estimate the
significance of the regression parameters. Significance threshold was set at p<0.05.

P á g i n a | 5 1
2.4.2. Regional GM and WM differences between cocaine-dependent individuals and non-
drug using controls
The general linear model implemented in SPM8 was used to conduct voxel-wise
comparisons between cocaine-dependent individuals and controls, and to determine the
relationship between consume severity factors (i.e. amount, duration and age of onset)
and regional GM and WM volume variations in cocaine-dependent individuals. In both
analyses the total volume of GM, or WM, was modeled as a linear confound in order to
account for residual global volume variability. The significance threshold was set at p<0.05
after family-wise correction for multiple comparisons (pFWE<0.05). Significant peaks
from the t-test maps are given in MNI space.
2.4.3. Regional relationships between GM and WM and measures of impulsivity and
estimates of drug use
We performed SPM8 multiple regression analysis, in which we tested the
significances of the Group (cocaine-dependent individuals vs. controls) x UPPS-P
dimensions interactions across the entire brain. We only included the UPPS-P dimensions
showing significant differences between the groups. The differences in the regression
coefficients as a function of group were tested using a significance threshold of p<0.05
after family-wise correction for multiple comparisons (pFWE<0.05).
3. Results
3.1. Participants’ characteristics
Both groups were matched on gender, ethnicity and language, and had statistically
equivalent distributions for age. Controls, compared to cocaine-dependent individuals, had
more years of education [(M=17.58, SD=4.56) vs. (M=12.03, SD=3.62)]; but analysis
including education as a covariate did not alter the results. Cocaine-dependent individuals
had greater scores for depression [(M=5.08, SD=3.98) vs. (M=1.32, SD=2.15)] and anxiety

P á g i n a | 5 2
[(M=7.05, SD=6.40) vs. (M=1.29, SD=2.29)], but on average both groups were below cut-
offs for clinical significance.
3.2. Trait Impulsivity
Cocaine-dependent individuals had higher scores than controls across the five
dimensions of the UPPS-P Scale. T-tests showed these differences were significant for lack
of premeditation (t=2.46, p=0.016, Cohen´s d=0.6), lack of perseverance (t=2.56, p=0.013,
Cohen´s d=0.6), negative urgency (t=5.14, p=0.000, Cohen´s d=1.2), and positive urgency
(t=5.86, p=0.000, Cohen´s d=1.4). We found no significant differences for sensation
seeking, such that this dimension was not further used in the subsequent analysis.
Table 2 Descriptive scores –means and standard deviations (in parentheses) of cocaine
users and non-drug using controls in the five UPPS-P dimensions.
UPPS-P Dimensions Cocaine-dependent
individuals Non-drug using
Controls Test
Negative Urgency 31.45 (6.79) 23.16 (6.99) (t46 = 5.14, p = 0.000)
Positive Urgency 32.91 (8.79) 21.66 (7.63) (t46 = 5.86, p = 0.000)
Lack of Premeditation 23.83 (5.95) 21 (3.68) (t46 = 2.46, p = 0.016)
Lack of Perseverance 21.26 (4.42) 18.84 (3.63) (t46 = 2.56, p = 0.013)
Sensation Seeking 30.63 (8.73) 29.89 (7.37) (t46 = 0,34, p = 0.73)
3.3. Imaging analysis
3.3.1. Global effects
For cocaine-dependent individuals, the TGMV/TBV ratio variability was accounted
for by monthly cocaine intake during regular use, t(34)=2.19, p=0.035, and age of onset,
t(34)=-2.33, p=0.026; the effect of duration of cocaine use also showed a trend to
significance, t(34)=-1.80, p=0.080. These variables explained about a 32% of the

P á g i n a | 5 3
variability in TGMV/TBV ratios, F(3,34)=5.21, p=0.005. Noteworthy, higher amounts of
cocaine (r=0.41, p=.01) and younger age of onset of cocaine use (r=-.34, p=0.04) were
associated with larger TGMV/TBV. The first result is associated with a significant decrease
in WM volumes (TWMV-amount of cocaine: r=0.43, p=0.006), but the second showed a
trend in relation to reductions in GM (TGMV-age of onset of cocaine abuse: r=-0.18, NS).
3.3.2. Regional GM and WM differences between cocaine-dependent individuals and non-
drug using controls
The cocaine group had significantly lower GM and WM volumes than controls.
Cocaine-dependent individuals showed lower GM volumes in a number of sections of the
OFC (left superior and medial frontal gyrus), right inferior frontal gyrus, right insula, left
amygdala and parahippocampal gyrus, left inferior and middle temporal gyrus, and
bilateral caudate (Table 3 and Figure 1). Likewise, cocaine dependent individuals showed
lower WM volumes in the left inferior and medial frontal gyrus, superior temporal gyrus,
right anterior cingulate cortex, insula and caudate (Table 4 and Figure 2).

P á g i n a | 5 4
Table 3 Summary of the results obtained by the gray matter voxel-wise analysis of volume reductions in cocaine-dependent individuals and non-
drug using controls.
Anatomical Region k Talairach Labels for peaks BA T p x y z
Frontal Lobe 3342 L Medial Frontal Gyrus Cingulate Gyrus
11, 10 32, 25, 24
5.81 5.75
0.004 0.005
-9 27 -13
Frontal Lobe Caudate
2802 R Inferior Frontal Gyrus R Caudate Head 47, 11, 34
6.39 5.53
0.000 0.010
28 9
21 19
-15 -15
Caudate 1118 L Caudate Head 5.52 0.010 6 -1 3 Limbic Lobe / Parahippocampal Gyrus
972 L Amygdala/Hippocampus 6.42 0.000 -33 -7 -19
Sub-Lobar 581 R Insula 13 5.81 0.004 52 -19 23
Temporal Lobe 3095 L Middle/Inferior Temporal Gyrus 21, 22, 20 5.98 0.002 -56 -14 -18

P á g i n a | 5 5
Figure 1 Brain regions showing significant gray matter decreases in cocaine-dependent
individuals with respect to non-drug using controls.
Table 4 Summary of the results obtained by the white matter voxel-wise analysis of
volume reductions in cocaine-dependent individuals and non-drug using controls.
Anatomical Region k Talairach Labels for peaks T p x y z
Frontal Lobe 1181 L Inferior Frontal Gyrus Sub-Gyral
6.07 0.001 -26 18 -17
Frontal Lobe 623 L Medial Frontal Gyrus 5.46 0.003 -20 57 -3
Temporal Lobe 389 L Superior Temporal Gyrus 5.24 0.007 -48 -25 -3
Frontal Lobe 1651 R Anterior Cingulate R Medial Frontal Gyrus
6.81 0.001 7 33 -15
Sub-Lobar 1531 Caudate Extra-Nuclear Lentiform Nucleus
5.86 0.001 12 12 -3
Temporal Lobe 674 Insula R Superior Temporal Gyrus R Transverse Temporal Gyrus
5.82 0.001 46 -26 12
Figure 2 Brain regions showing significant white matter decreases in cocaine-dependent
individuals with respect to non-drug using controls.

P á g i n a | 5 6
3.3.3. Regional relationships between GM and WM and measures of impulsivity and
estimates of drug use
We found a number of regions in which GM volumes showed differential
associations with the impulsivity dimensions of lack of premeditation and negative
urgency; in all cases, there was a significant Group x Impulsivity interaction (see Table 5).
The associations between impulsivity levels and GM volumes were significantly different
for cocaine-dependent individuals vs. controls in the left inferior frontal gyrus, right insula
and left putamen (in relation to lack of premeditation), and left middle frontal gyrus and
sub-gyral (in relation to negative urgency). Left inferior frontal gyrus was positively
associated with lack of premeditation in cocaine users, whereas it was negatively
correlated with this dimension in controls. Conversely, right insula and left putamen were
negatively correlated with lack of premeditation in cocaine users, but did not correlate
with this dimension in controls. With regard to negative urgency, the subgyral showed
significant positive correlations in the cocaine group, whereas the association was
negative in controls. Finally, in the case of the left middle frontal gyrus, none of the slopes
significantly differed from zero, but the trend was positive in cocaine-dependent
individuals, and negative in controls.
We did not find significant associations between WM volumes and the different
dimensions of impulsivity in the cocaine or control groups. Finally, we did not find
significant associations between the main patterns of drug use (amount, duration and age
of onset of cocaine abuse) and GM and WM volumes.

P á g i n a | 5 7
Table 5 Regression slopes showing different associations between impulsivity and gray matter volumes between cocaine-dependent individuals
(CDI) and non-drug-using controls (NDC) (pFWE=0.05).
Anatomical Region k Talairach Label BA p (β) Slopes Volume x y z
Lack of premeditation
Sub-lobar 179 R Insula BA 47/BA 13 0.02 CDI-/NDC+ CDI<NDC 30 10 -3
358 L Putamen 0.05 CDI-/NDC+ CDI<NDC -21 10 -4
Fontal Lobe 249 L Inferior Frontal Gyrus BA 10 0.005 CDI+/NDC- CDI<NDC -37 41 1
Negative Urgency
Frontal Lobe 228 L Middle Frontal Gyrus BA10/46 0.038 CDI+/NDC- CDI<NDC -39 37 19
381 R Sub-Gyral BA 8 0.024 CDI+/NDC- CDI<NDC 25 24 36

P á g i n a | 5 8
4. Discussion
Our results showed that cocaine-dependent individuals, compared to controls,
have lower GM volumes in the OFC cortex, anterior cingulate, inferior frontal gyrus, insula,
amygdala, temporal gyrus, and caudate. We also found WM reductions in adjacent regions
of the anterior cingulate, inferior/middle frontal gyrus, insula and putamen. As expected,
we found significant differences in the way that impulsivity dimensions correlates with
GM volumes in cocaine users vs. controls: cocaine patients had elevated levels of trait
impulsivity, and these scores were differentially associated with GM volumes in the left
inferior frontal gyrus, insula and putamen (lack of premeditation), and left middle frontal
gyrus (negative urgency). Inferior and medial superior frontal clusters positively
correlated with impulsivity in cocaine users, and negatively in controls; conversely, insula
and putamen correlated negatively with impulsivity in cocaine patients and showed no
correlations in controls. These results point to a distinctive link between trait impulsivity
and regional GM among cocaine-dependent individuals, which specifically relates to
frontostriatal systems regions. The measures of patterns of cocaine use correlated with
general indices of GM/TBV ratios but had no significant influence on GM volumes at the
stricter voxel level. Overall, our findings are in agreement with the notion that GM
reductions in cocaine-dependent individuals are at least partly mediated by trait
impulsivity.
Our findings of GM reductions in cocaine-dependent individuals are largely
consistent with previous neuroimaging studies in revealing structural abnormalities in
brain regions relevant to reinforcement learning (amygdala, OFC) and inhibitory control
(anterior cingulate, inferior frontal gyrus, insula and caudate) (Ersche et al., 2011;
Franklin et al., 2002; Makris et al., 2004; Matochik et al., 2003). Recent findings from fMRI
connectivity analyses also provide support to the notion that neural networks connecting
the medial frontal cortex with paralimbic and temporal regions are significantly less

P á g i n a | 5 9
functional in cocaine users (Gu et al., 2010). Biological abnormalities in these regions are
thought to underlie psychological alterations leading to overestimation of the motivational
relevance of drug-related reinforcement (the impulsive system) and to breakdown of
inhibitory processes necessary to achieve long-term abstinence (the reflective system)
(Bechara, 2005; Goldstein and Volkow, 2002; Redish et al., 2008; Verdejo-García and
Bechara, 2009). These alterations are proposed to anchor pervasive drug taking in the face
of increasing aversive consequences, one of the key features of addiction (DSM-IV).
A key unresolved issue is that if these structural abnormalities are reflective of
cocaine induced brain attrition, or are related to personality traits, or reflect a
combination of both factors. Our findings, in agreement with growing neuroscience
evidence, are supportive of the impact of personality traits. However, these traits may
interact with cocaine use in producing GM alterations, as suggested by the global TGMV
analysis. Recent studies indicate that, in healthy individuals, impulsivity is negatively
correlated with midbrain and striatal D2/D3 receptors availability (Buckholtz et al., 2010;
Lee et al., 2009), but this association is stronger in methamphetamine dependent
individuals (Lee et al., 2009) and correlates with amphetamine-induced dopamine release
in the striatum (Buckholtz et al., 2010). Striatal D2 availability is associated with metabolic
rate in the prefrontal cortex of psychostimulant users (Volkow et al., 2001). Therefore,
different strands of evidence indicate that frontostriatal systems mediate impulsive
behavior and thereby promote psychostimulant addiction. Nonetheless, we acknowledge
that these issues can only be tested by longitudinal imaging studies in individuals at high
risk of developing psychostimulant dependence, or by gene association imaging studies
(Kreek et al., 2005; Verdejo-García et al., 2008).
In accordance with the notion that impulsivity is a multifaceted construct (Smith et
al., 2007; Whiteside and Lynam, 2001), our results showed differential links between
impulsivity dimensions and GM volumes. Lack of premeditation was positively associated

P á g i n a | 6 0
with left inferior frontal gyrus GM in cocaine users but not controls. This region is
importantly involved in the representation of expected values (Bermpohl et al., 2010);
such that persistent stimulation by drug-expected rewards could have induced GM
enlargements linked to an increased tendency to rapidly engage in non-adequately
forethought behavior. On the other hand, both insula and putamen have been involved in
drug-related neuroadaptations related to attentional bias and craving (Childress et al.,
2008; Luijten et al., 2011; Volkow et al., 2006), such that reduced volumes may predict
increased prepotency of drug-related action tendencies. On the other hand, negative
urgency was positively associated with GM in subgyral BA 8 in cocaine users, negatively in
controls. This region has been previously associated with the experience of uncertainty
(Volz et al., 2003), which is a key aspect of the associative learning processes mediating
expected drug effects (D’Souza and Duvauchelle, 2008). In fact, adjacent regions of the
medial frontal gyrus (BA 10) display increased activation during uncertain outcome
selection in psychostimulant users (Leland et al., 2006). Therefore, in this case the
rationale could be similar to that posited for inferior frontal gyrus; repeated exposure to
uncertainty-loaded scenarios may be associated with increased subgyral volumes and
higher risk to trigger impulsive acts when under emotionally uneasy conditions.
Strengths of this study include the adequate sample size, the sound clinical
characterization and the agreement between results from the different analytical
approaches (between-group differences and regressions testing the link between
impulsivity and GM). On the other hand, we should also acknowledge a number of
limitations. The cross-sectional design precludes us from drawing conclusions about the
causality of GM deficits; one way out of addressing this issue would be by using
longitudinal designs, but these studies are costly and convey important ethical concerns
and methodological complexities. Furthermore, we cannot completely rule out the
influence of depression and anxiety levels, although they were on average below the cut-
off for clinical significance and they had very low impact on regression models when

P á g i n a | 6 1
included. Finally, although we discuss our results as pertaining to cocaine dependence,
polysubstance abuse is almost ubiquitous among cocaine-dependent individuals.
However, the current sample was carefully selected for relative specificity of cocaine
related problems and low use of other drugs. The illegal drugs more frequently and
intensely co-abused in the sample were cannabis and MDMA, but estimates of the use of
these drugs failed to predict regional GM.

P á g i n a | 6 2
References
Alia-Klein, N., Parvaz, M.A., Woicik, P.A., Konova, A.B., Maloney, T., Shumay, E., et al., 2011.
Gene x disease interaction on orbitofrontal gray matter in cocaine addiction. Arch Gen
Psychiatry 68, 283-294.
Ashburner, J., Friston, K.J., 2005. Unified segmentation. Neuroimage 26, 839-851.
Barrós-Loscertales, A., Garavan, H., Bustamante, J.C., Ventura-Campos, N., Llopis, J.J.,
Belloch, V., et al., 2011. Reduced striatal volume in cocaine-dependent patients.
Neuroimage 56, 1021-1026.
Bechara, A., 2005. Decision making, impulse control and loss of willpower to resist drugs:
a neurocognitive perspective. Nat Neurosci. 8, 1458-1463.
Belin, D., Mar, A.C., Dalley, J.W., Robbins, T.W., Everitt, B.J., 2008. High impulsivity predicts
the switch to compulsive cocaine-taking. Science 320, 1352-1355.
Bermpohl, F., Kahnt, T., Dalanay, U., Hägele, C., Sajonz, B., Wegner, T., et al., 2010. Altered
representation of expected value in the orbitofrontal cortex in mania. Hum Brain Mapp.
31, 958-969.
Buckholtz, J.W., Treadway, M.T., Cowan, R.L., Woodward, N.D., Li, R., Ansari, M.S., et al.,
2010. Dopaminergic network differences in human impulsivity. Science 329, 532.
Childress, A.R., Ehrman, R.N., Wang, Z., Li, Y., Sciortino, N., Hakun, J., et al., 2008. Prelude to
passion: limbic activation by "unseen" drug and sexual cues. Plos One 3, e1506.
Cyders, M.A., Smith, G.T., Spillane, N.S., Fischer, S., Annus, A.M., Peterson, C., 2007.
Integration of impulsivity and positive mood to predict risky behavior: development and
validation of a measure of positive urgency. Psychol Assess. 19, 107-118.

P á g i n a | 6 3
Dalley, J.W., Fryer, T.D., Brichard, L., Robinson, E.S., Theobald, D.E., Lääne, K., et al., 2007.
Nucleus accumbens D2/3 receptors predict trait impulsivity and cocaine reinforcement.
Science 315, 1267-1270.
D'Souza, M.S., Duvauchelle, C.L., 2008. Certain or uncertain cocaine expectations influence
accumbens dopamine responses to self-administered cocaine and non-rewarded operant
behavior. Eur Neuropsychopharmacol. 18, 628-638.
EMCDDA European Monitoring Centre for Drugs and Drug Addiction (2009): Annual
report 2009: the state of the drugs problem in Europe Luxembourg. Publications Office of
the European Union.
Ersche, K.D., Barnes, A., Simon Jones, P., Morein-Zamir, S., Robbins, T.W., Bullmore, E.T.,
2011. Abnormal structure of frontostriatal brain systems is associated with aspects of
impulsivity and compulsivity in cocaine dependence. Brain 134, 2013-2024.
Ersche, K.D., Bullmore, E.T., Craig, K.J., Shabbir, S.S., Abbott, S., Müller, U., et al., 2010.
Influence of compulsivity of drug abuse on dopaminergic modulation of attentional bias in
stimulant dependence. Arch Gen Psychiatry 67, 632-644.
Franklin, T.R., Acton, P.D., Maldjian, J.A., Gray, J.D., Croft, J.R., Dackis, C.A., et al., 2002.
Decreased gray matter concentration in the insular, orbitofrontal, cingulate, and temporal
cortices of cocaine patients. Biol Psychiatry 51, 134-142.
Goldstein, R.Z., Tomasi, D., Alia-Klein, N., Zhang, L., Telang, F., Volkow, N.D., 2007. The
effect of practice on a sustained attention task in cocaine abusers. Neuroimage 35, 194-
206.
Goldstein, R.Z., Volkow, N.D., 2002. Drug addiction and its underlying neurobiological
basis: neuroimaging evidence for the involvement of the frontal cortex. Am J Psychiatry
159, 1642-1652.

P á g i n a | 6 4
Gu, H., Salmeron, B.J., Ross, T.J., Geng, X., Zhan, W., Stein, E.A., et al., 2010.
Mesocorticolimbic circuits are impaired in chronic cocaine users as demonstrated by
resting-state functional connectivity. Neuroimage 53, 593-601.
Koob, G.F., Volkow, N.D., 2010. Neurocircuitry of addiction. Neuropsychopharmacology 35,
217-238.
Kreek, M.J., Bart, G., Lilly, C., LaForge, K.S., Nielsen, D.A., 2005. Pharmacogenetics and
human molecular genetics of opiate and cocaine addictions and their treatments.
Pharmacol Rev. 57, 1-26.
Lee, B., London, E.D., Poldrack, R.A., Farahi, J., Nacca, A., Monterosso, J.R., et al., 2009.
Striatal dopamine d2/d3 receptor availability is reduced in methamphetamine
dependence and is linked to impulsivity. J Neurosci. 29, 14734-14740.
Leland, D.S., Arce, E., Feinstein, J.S., Paulus, M.P. 2006. Young adult stimulant users'
increased striatal activation during uncertainty is related to impulsivity. Neuroimage 33,
725-731.
Luijten, M., Veltman, D.J., van den Brink, W., Hester, R., Field, M., Smits, M., et al., 2011.
Neurobiological substrate of smoking-related attentional bias. Neuroimage 54, 2374-2381.
Makris, N., Gasic, G.P., Seidman, L.J., Goldstein, J.M., Gastfriend, D.R., Elman, I., et al., 2004.
Decreased absolute amygdala volume in cocaine addicts. Neuron 44, 729-740.
Matochik, J.A., London, E.D., Eldreth, D.A., Cadet, J.L., Bolla, K.I., 2003. Frontal cortical tissue
composition in abstinent cocaine abusers: a magnetic resonance imaging study.
Neuroimage 19, 1095-1102.
Matsuo, K., Nicoletti, M., Nemoto, K., Hatch, J.P., Peluso, M.A., Nery, F.G., et al., 2009. A voxel-
based morphometry study of frontal gray matter correlates of impulsivity. Hum Brain
Mapp. 30, 1188-1195.

P á g i n a | 6 5
Narayana, P.A., Datta, S., Tao, G., Steinberg, J.L., Moeller, F.G., 2010. Effect of cocaine on
structural changes in brain: MRI volumetry using tensor-based morphometry. Drug
Alcohol Depend. 111, 191-199.
Redish, A.D., Jensen, S., Johnson, A., 2008. A unified framework for addiction:
vulnerabilities in the decision process. Behav Brain Sci. 31, 415-437.
Sim, M.E., Lyoo, I.K., Streeter, C.C., Covell, J., Sarid-Segal, O., Ciraulo, D.A., et al., 2007.
Cerebellar gray matter volume correlates with duration of cocaine use in cocaine-
dependent subjects. Neuropsychopharmacology 32, 2229-2237.
Smith, G.T., Fischer, S., Cyders, M.A., Annus, A.M., Spillane, N.S., 2007. On the validity and
utility of discriminating among impulsivity-like traits. Assessment 14, 155-170.
Substance Abuse and Mental Health Services Administration, 2010. Results from the 2009
National Survey on Drug Use and Health: Mental Health Findings (Office of Applied
Studies, NSDUH Series H-39, HHS Publication No. SMA 10-4609). Rockville, MD.
Tanabe, J., Tregellas, J.R., Dalwani, M., Thompson, L., Owens, E., Crowley, T., et al., 2009.
Medial orbitofrontal cortex gray matter is reduced in abstinent substance-dependent
individuals. Biol Psychiatry 65, 160-164.
Verdejo-García, A., Bechara, A., 2009. A somatic marker theory of addiction.
Neuropharmacology 56, 48-62.
Verdejo-García, A., Lawrence, A.J., Clark, L., 2008. Impulsivity as a vulnerability marker for
substance-use disorders: review of findings from high-risk research, problem gamblers
and genetic association studies. Neurosci Biobehav Rev. 32, 777-810.
Verdejo-García, A., Lozano, O., Moya, M., Alcázar, M.A., Pérez-García, M., 2010.
Psychometric properties of a Spanish version of the UPPS-P impulsive behavior scale:

P á g i n a | 6 6
reliability, validity and association with trait and cognitive impulsivity. J Pers Assess. 92,
1532-7752.
Verdejo-García, A.J., López-Torrecillas, F., Aguilar de Arcos, F., Pérez-García, M., 2005.
Differential effects of MDMA, cocaine, and cannabis use severity on distinctive components
of the executive functions in polysubstance users: a multiple regression analysis. Addict
Behav. 30, 89-101.
Volkow, N.D., Wang, G.J., Telang, F., Fowler, J.S., Logan, J., Childress, A.R. et al., 2006.
Cocaine cues and dopamine in dorsal striatum: mechanism of craving in cocaine addiction.
J Neurosci. 26, 6583-6588.
Volkow, N.D., Chang, L., Wang, G.J., Fowler, J.S., Ding, Y.S., Sedler, M., et al., 2001. Low level
of brain dopamine D2 receptors in methamphetamine abusers: association with
metabolism in the orbitofrontal cortex. Am J Psychiatry 158, 2015-2021.
Volz, K.G., Schubotz, R.I., von Cramon, D.Y. 2003. Predicting events of varying probability:
uncertainty investigated by fMRI. Neuroimage 19, 271-280.
Whiteside, S.P., Lynam, D.R., 2001. The five factor model and impulsivity: using a structural
model of personality to understand impulsivity. Pers Individ Dif. 30, 669-689.

P á g i n a | 6 7
* ARTÍCULO 2
Neural Correlates of the Severity of Cocaine, Heroin, Alcohol, MDMA and Cannabis
Use in Polysubstance Abusers: A Resting-PET Brain Metabolism Study
* Moreno-López, L., Stamatakis, E. A., Fernández-Serrano, M. J., Gómez-Río, M., Rodríguez-
Fernández, A., Pérez-García, M., & Verdejo-García, A. (2012). Neural Correlates of the
Severity of Cocaine, Heroin, Alcohol, MDMA and Cannabis Use in Polysubstance Abusers: A
Resting-PET Brain Metabolism Study. PLoS ONE, en prensa.

P á g i n a | 6 8
1. Introduction
Drug addiction has been associated with neuroadaptations in brain systems
involved in motivation, memory and executive control [1]. Functional neuroimaging
studies have revealed that the use of different classes of drugs is associated with
dysfunctions in a range of overlapping brain regions including ventromedial and
dorsolateral prefrontal cortex (DLPFC), anterior cingulate cortex, inferior frontal gyrus,
insula, amygdala, basal ganglia and cerebellum [2]. These dysfunctions may explain the
overlapping cognitive deficits observed in users of different drugs, including working
memory, inhibitory control, flexibility or decision-making deficits [3]. In addition, some
specific effects have been described for the association between cocaine use and the
parietal cortex [4,5,6], heroin use and the temporal cortex [7], MDMA use and occipital,
hippocampus and thalamic regions [8,9], and cannabis use and the premotor cortex
[10,11]. These associations are congruent with relatively specific neuropsychological
deficits pertaining to attention and cognitive control in cocaine users, long-term memory
in opiate users, visuospatial memory in MDMA users, and psychomotor function in
cannabis users [3,7]. In spite of the available evidence, in vivo studies have not been
systematically conducted to look at associations between severity of use of different drugs
and brain dysfunction. In fact, several neuroimaging studies have failed to detect such link
or have provided controversial results (reviews in 12). These negative findings and
controversies might be linked to the impact of relevant confounding variables. One of
these is the fact that neuroimaging studies have frequently tested drug users currently
using drugs or having brief periods of abstinence (24-48 h) [13, 14]; under these
conditions, the presumed link between lifetime drug use and brain dysfunction could be
masked by several other factors, including recent drug use or psychological symptoms
associated with withdrawal and short-term abstinence [2]. Another key variable for
consideration is the concurrent use of multiple types of substances, which can introduce
significant confounds in the interpretation of the data.

P á g i n a | 6 9
The current study aimed to specifically address the association between lifetime
estimates of use of different types of drugs and brain functioning (as measured by 18F-
fluorodeoxyglucose Positron Emission Tomography (FDG-PET), statistically controlling for
the concurrent use of other drugs in a sample of polysubstance users with prolonged
abstinence from all drugs. Specifically, we examined the link between lifetime estimates of
amount and duration of use of cocaine, heroin, alcohol, MDMA and cannabis and brain
metabolism (BM). Based on previous findings we hypothesized that exposure to all
substances would be significantly associated with overlapping reductions of metabolism
in frontal, limbic, striatal and cerebellar regions, whereas cocaine and heroin use would
specifically correlate with parietal and temporal cortices respectively.
2. Methods
2.1 Participants
Forty-nine substance dependent individuals (SDI), forty-one men and eight women
with a mean age of 32.67 years, were recruited during their stay in an inpatient treatment
program at the centre “Proyecto Hombre” in Granada (Spain). This centre provides a
controlled setting for the treatment of substance use disorders. Selection criteria for
participants in this study were (i) meeting the DSM-IV criteria for substance dependence;
(ii) absence of documented comorbid mood or personality disorders, as assessed by
clinical reports; (iii) absence of documented head injury or neurological disorders; and
(iv) to have minimum abstinence duration from all drugs consumed (except for tobacco)
of 15 days before testing. In fact, SDI participants had overall much longer abstinence
duration (mean of 32.94, SD=11.25 weeks); for that reason it was possible to rule out the
presence of withdrawal symptoms or brain function alterations associated with the acute
or short term effects of the drugs. None of the participants were experiencing withdrawal
symptoms as assessed by routine medical examination or were enrolled in opioid

P á g i n a | 7 0
substitution treatments with methadone or other pharmacological treatments (e.g.,
benzodiazepines) during the course of the assessment interview/PET evaluations.
2.2 Testing protocols and procedures
This study was approved by the Human Subjects Committee of the University of
Granada. One-step Syva urine drug screens for amphetamines, benzodiazepines, cannabis,
cocaine and opiates were conducted to confirm abstinence at time of testing. Participants
were evaluated individually in two different sessions. During the first session, the
participants were provided with information about the study and were encouraged to ask
questions to ascertain they fully understood the rational for the study and were clear on
the details for participation. The participants then provided written consent and were
assessed with the Interview for Research on Addictive Behaviours [15] in the “Proyecto
Hombre” installations”. During the second session, each patient was accompanied by a
therapist to the nuclear medicine service of the “Virgen de las Nieves” hospital, where the
PET neuroimaging session took place. This second session usually took place one week
after the first session was completed.
2.3 Tools
2.3.1 Drug Use Information
The Interview for Research on Addictive Behaviour [15] was used to examine the
severity of drug use. This interview evaluates, by means of a brief interview, the quantity
(average dosing), frequency (number of drug taking episodes per month), and duration
(years of duration) of the use of different substances that can produce physical or
psychological dependence, including cocaine, heroin, alcohol, MDMA and cannabis which
were the main drugs of choice in the present study. For every substance the participant
had actually used, the following information was requested: (i) the average dose of each
target drug taken in each episode of use (number of grams for cocaine and heroin, number

P á g i n a | 7 1
of units for alcohol, considering that 25 cl. of hard liquor (e.g., scotch) equals 2 units, while
25 cl. of wine or beer equals 1 unit, number of pills for MDMA and number of cigarettes for
cannabis); (ii) the frequency of these consumption episodes per month (daily, between
one and three times upon a week, once a week, between one and three times upon a
month, or once a month); and (iii) the number of years that elapsed since the onset of use.
From this information, we obtained two indices for each of the drugs used: (i) Amount
(average dose x frequency), an index of monthly use of the drug; and (ii) Duration of drug
use, measuring number of years of exposure to the drug. The main sociodemographic and
clinical features from the sample are displayed in Table 1.
Table 1 Sociodemographic data and clinical features of the participants
Subjects Characteristics Gender Male 41
Female 8
Mean S.D. Maximum Minimum
Age (years) 32.67 8.03 53 21
Education (years) 9.73 2.34 17 5
Amount of drug usea Substance Mean S.D. Maximum Minimum Cocaine (g) (46) 48.69 44.83 180 0
Heroin (g) (16) 9.12 20.63 120 0
Alcohol (units) (45) 571.24 489.34 1800 0
MDMA (pills) (23) 13.41 24.31 120 0
Cannabis (cigarettes) (37) 148.57 190.64 750 0
Duration of drug use (years) Substance Mean S.D. Maximum Minimum Cocaine 7.95 5.95 23 0
Heroin 1.76 3.69 17 0
Alcohol 10.85 7.67 27 0
MDMA 1.40 2.28 8 0
Cannabis 8.31 8.13 29 0
Abstinence (weeks) 32.94 11.25 60 12 a Amount of monthly use of the drug calculated using the average dose (per episode) × frequency (per month).

P á g i n a | 7 2
2.3.2 PET Image Acquisition
PET scans were acquired with an ECAT/931 scanner (Siemens CTI ECAT EXACT),
at the service of nuclear medicine centre of the hospital “Virgen de las Nieves” in Granada
(Spain). For each individual, we obtained 20 minute emission scans 30 minutes after the
injection of one dose of 200 MBq of FDG administrated only after levels of glycemia had
been checked (they must be below 120 mg/dl) [16]. The subjects were scanned with their
eyes open and ears unplugged in a dimly illuminated quiet room. Raw data were
processed using an iterative reconstruction technique (OSEM method: 10 iterations, 32
subsets). Images were reoriented in transaxial, coronal and saggital planes. For the
analyses reported here the reconstructed images used had 47 axial slices each with an in-
plane resolution of 2.57x2.57 and a slice thickness of 3.38mm.
2.4 Data analysis
2.4.1 Preprocessing of PET images
PET images were converted from DICOM to NIFTI format and were spatially
normalized to the SPM PET template (Wellcome Department of Cognitive Neurology,
London, UK) using linear (12 affine) and non-linear normalization. An average image of all
the normalized images was obtained and this acted as a study specific PET template. In
this manner we constructed a template in the same modality, from the same scanner as
the images we wanted to normalize. The raw PET images were then spatially normalized
with linear affine and nonlinear parameters to this study-specific template. Visual
inspection revealed this optimized spatial normalization technique produced satisfactory
spatial normalization for the PET image of every volunteer.
2.4.2 Statistical Analysis

P á g i n a | 7 3
The spatially normalized PET images were smoothed with an 8mm3 isotropic
Gaussian filter and were statistically modeled using the General Linear Model in SPM5.
Linear regressions were used to carry out several analyses in order to identify brain areas
with a significant correlation between uptake values and measures of the amount and
duration of cocaine, heroin, alcohol, MDMA and cannabis use. Not all subjects used all the
drugs but since virtually all participants had regularly abused two or more substances, the
analyses were conducted on the whole group and results were interpreted in relation to
polysubstance use. In each of the analysis, we controlled for the effects of the other
substances concurrently used (e.g., analysis of severity of cocaine use controlled for the
effects of co-abuse of heroin, alcohol, MDMA and cannabis), age and years of education.
We report local maximum peaks that survive a voxel threshold of p<0.001 (uncorrected
for multiple comparisons, cluster size ≥100 voxels). We chose to report our findings at this
threshold based on numerous similar studies of PET imaging in drug addiction [17,18].
3. Results
The main results are summarized in Table 2.
3.1 Amount of use
Voxel-based analyses showed negative correlations between lifetime amount of
cocaine, alcohol and cannabis use and several brain regions. The amount of cocaine used
was negatively correlated with a cluster encompassing the left inferior parietal lobule
extending to the left postcentral gyrus (Figure 1.1). The amount of alcohol used was
negatively associated with three clusters: one cluster included the left middle and superior
temporal cortex; a second cluster included the bilateral superior frontal cortex extending
to the left DLPFC and the right supplementary motor area; and a third cluster included the
right DLPFC extending to the superior frontal gyrus (Figure 1.2). The amount of cannabis
use showed a negative correlation with two clusters: one cluster included the left inferior
frontal gyrus (pars triangularis) extending to the DLPFC, and a second cluster included the

P á g i n a | 7 4
right DLPFC extending to the superior frontal cortex (Figure 1.3). We did not find
significant correlations between the amount of heroin or MDMA use and BM measures.
3.2 Duration of use
Voxel-based analyses showed negative correlations between duration of heroin,
alcohol and MDMA use and several brain regions. Duration of heroin use showed a
negative correlation with three clusters, which encompassed the right inferior, middle and
superior temporal cortex extending to the right supramarginal and inferior frontal gyrus
(Figure 1.4). Duration of alcohol use was negatively correlated with four clusters. One
cluster included the right inferior temporal cortex extending to the middle aspect of this
area. A second cluster included the right putamen and pallidum. A third cluster
encompassed the right precentral and postcentral gyrus and the fourth cluster included
the right fusiform and parahippocampal gyrus (Figure 1.5). Finally, we found negative
correlations between the duration of MDMA use and BM in three clusters: the first cluster
included the left postcentral and inferior parietal gyrus; the second cluster included the
right inferior frontal gyrus (pars triangularis) extending to the DLFPC; and the third
cluster included the right superior temporal pole extending to the middle part of this
region (Figure 1.6).

P á g i n a | 7 5
Table 2 Relation between severity of drug use and PET uptake.
Substance Voxels-Based
Analysis
BA KE T MNI
Amount
Cocaine Parietal Inf L 40 176 3.98 -58 -38 54
Alcohol Temporal Mid L 21 267 4.04 -66 -2 -10
Frontal Sup L 9 308 3.96 -20 30 36
Frontal Sup R 6 365 3.73 22 2 60
Frontal Mid R 46 372 3.82 26 22 34
Cannabis Frontal Inf Tri L 45 355 4.40 -44 22 30
Frontal Mid R 46 125 4.02 24 48 28
Duration
Heroin Frontal Mid R 46 876 3.77 44 36 34
Temporal Inf R 20 257 4.58 62 -16 -32
Temporal Sup R 22 1090 4.19 66 -44 20
Alcohol Temporal Inf R 20 238 4.22 64 -16 -30
Putamen R 472 3.71 20 -2 8
Precentral R 6 274 3.67 50 2 34
Fusiform R 20 299 3.65 30 -6 -40
MDMA Postcentral L 40 131 3.73 -28 -40 48
Frontal Inf Tri R 45 139 3.67 44 34 20
Temporal Pole Sup R 20 561 3.67 32 12 -30 Voxel-based analyses at p<0.001 (uncorrected, cluster size > 100 voxels). Labels were obtained using the aal toolbox (Tzourio-Mazoyer et al., 2002). Broadmann areas were obtained using MRICron (Rorden and Brett, 2000). KE indicate the number of voxels included in the cluster. Stereotaxic coordinades are those listed in SPM5. Inf, inferior; Sup, superior; L, left; R, right; Supp, supplementary; Mid, middle; Orb, orbitalis.

P á g i n a | 7 6
Figure 1. Areas of reduced metabolism predicted by the amount or duration of drug
use: (1) amount of cocaine, (2) amount of alcohol, (3) amount of cannabis (4) duration of
heroin (5) duration of alcohol and (6) duration of MDMA. Areas of reduced metabolism are
superimposed on a T1 weighted MRI image in MRICRON. MNI coordinates are shown
underneath each panel.

P á g i n a | 7 7
4. Discussion
Our findings showed that the estimates of severity of use of heroin, alcohol, MDMA
and cannabis were negatively associated with DLPFC and temporal cortex regional
metabolism in polysubstance dependent individuals with prolonged abstinence from all
these drugs. Alcohol use was further associated with lower metabolism in frontal
premotor cortex and putamen. Furthermore, severity of stimulants use (cocaine and
MDMA) was uniquely associated with inferior parietal/postcentral cortex metabolism.
These associations were established after a prolonged period of abstinence, and after
controlling for multiple confounding variables, thus surpassing the limitations of previous
functional neuroimaging studies that were not specifically designed to study prolonged
abstinence, and were confounded by important variables, such as withdrawal related
symptoms or concurrent use of multiple substances. The associations found are in
agreement with our initial predictions, and with the neuropsychological deficits typically
aligned with the chronic use of these substances; these include reduced competency in
executive functions (related to all classes of drugs), episodic memory (mainly observed on
heroin, alcohol, MDMA and cannabis users), motor control (alcohol and cannabis users),
and visuospatial attention (stimulants users) (see meta-analyses in 3,8,19,20).
As expected, the estimates of amount or duration of different classes of drugs were
negatively associated with overlapping regions within the DLPFC (BA 9, 45, 46). This
region has been linked with several of the key neuroadaptations associated with drug
addiction, including drug conditioning, loss of self-control, and stimulus-driven
compulsive behavior [2]. This finding is also in agreement with results from several
neuropsychological studies showing cognitive deficits on working memory, planning or
inhibition processes, which are associated with DLPFC functioning, across users of several
drugs [3]. We also found negative associations between estimates of duration of heroin,
alcohol, and MDMA use and regional metabolism on partially overlapping sections of the

P á g i n a | 7 8
temporal cortex. These associates may substantiate the links between the intensity and
duration of involvement with these drugs and the degree of memory attrition, which has
been observed in longitudinal studies with users of these substances [21,22]. It is worth
mentioning that duration of heroin use was shorter to that of cocaine and alcohol use;
however, previous brain structural findings indicate that duration of heroin use is a
critical factor leading to brain damage, even in users with periods of use below five years
[23]. Surprisingly, we failed to find significant associations between the severity of the
substances used and overlapping limbic, striatal and cerebellar regions but this result
could be explained from the fact that these regions play a greater role during current use
and short-term abstinence [24]. In addition, we found relatively specific negative
correlations between amount of cocaine use and regional metabolism in the inferior
parietal cortex, and between alcohol duration and regions involved in motor programming
and control (frontal precentral and putamen). The link between cocaine use and parietal
dysfunction has been observed on previous functional imaging studies during cognitive
challenges of attention and executive control [6,25,26]. The link between alcohol use and
precentral and basal ganglia dysfunction is in agreement with structural imaging findings
showing that function of these regions is significantly reduced in alcohol users compared
to other forms of addiction [27]. Additional evidence comes from volumetric studies which
associated volumetric measures of these regions and deficient motor skills in alcoholics
[28]. These more specific findings could be used to design tailored interventions aimed to
address specific aspects of frontal-subcortical executive functions in different profiles of
substance abusers.
An important implication of this study is the fact that the link between severity of
drug use and BM is still observable in substance abusers with prolonged periods of
abstinence (an average of eight months in this sample). Therefore, quantity and duration
of drug use may still be affect brain functioning after several months of successful
cessation of drug use. This is particularly relevant in view of recent findings indicating that

P á g i n a | 7 9
certain patterns of brain dysfunction can reliably predict mid-term and long-term relapse.
Permanent brain alterations may also have important repercussions for the rehabilitation
of SDI, since these alterations can affect the ability of SDI to assimilate the contents and
participate in the activities of the rehabilitation programs [29,30]. In fact, preliminary
evidence suggests that cognitive rehabilitation techniques, such as errorless learning may
be successful in compensating chronic cognitive deficits in substance abusers [31]. Thus,
the brain networks impacted by severity of drug abuse can play a central role in clinical
outcome and relapse and should become priority targets for therapeutic interventions.
Several possible limitations of our study should be considered and addressed by
future research. Our findings were not obtained in “pure” users of each of the substances
but in mostly polysubstance users. Although this limitation is inherent to the clinical
literature on addiction (i.e., “pure” users of one single drug are quite rare among substance
abusers enrolled in treatment programs), we attempted to control for the co-abuse of
other drugs using statistics. A related confounding variable was the use of nicotine, which
is ubiquitous among drug users but was not specifically assessed or controlled for.
Furthermore, due to the use of a correlational design, this study cannot yield conclusions
about cause-effect relationships between the use of drug and resting BM. However, given
the agreement of our results with both resting state PET, activation PET/fMRI and
addiction neuropsychology studies, we could hypothesize that, at least in part, the
alterations observed in this population could be due to the quantity and duration related
effects of the consumption of the substances under consideration in this study.
One last point to consider is that our findings could be due to premorbid brain
alterations or the results of the interaction between the premorbid alterations and the
neurotoxic effects of drug use. This question should be addressed through longitudinal
designs. A related limitation is that no comparison with a healthy control database was
performed. As we argued before, a healthy control group was not needed to test the main

P á g i n a | 8 0
predictions of our study; however, the lack of controls has stopped us from making any
assumption about the direction of these findings in relation to the healthy population.

P á g i n a | 8 1
References
1. Koob GF, Volkow ND (2010) Neurocircuitry of addiction. Neuropsychopharmacology 35:
217-238.
2. Goldstein RZ, Volkow ND (2011) Dysfunction of the prefrontal cortex in addiction:
neuroimaging findings and clinical implications. Nat Rev Neurosci 12: 652-669.
3. Fernández-Serrano MJ, Pérez-García M, Verdejo-García A (2011) What are the specific
vs. generalized effects of drugs of abuse on neuropsychological performance? Neurosci
Biobehav Rev 35: 377-406.
4. Aron JL, Paulus MP (2007) Location, location: using functional magnetic resonance
imaging to pinpoint brain differences relevant to stimulant use. Addiction 1: 33-43.
5. Lane SD, Steinberg JL, Ma L, Hasan KM, Kramer LA, et al. (2010) Diffusion tensor
imaging and decision making in cocaine dependence. PLoS One 5: e11591.
6. Tomasi D, Goldstein RZ, Telang F, Maloney T, Alia-Klein N, et al. (2007) Widespread
disruption in brain activation patterns to a working memory task during cocaine
abstinence. Brain Res 1171: 83-92.
7. Gruber SA, Silveri MM, Yurgelun-Todd DA (2007) Neuropsychological consequences of
opiate use. Neuropsychol Rev 17: 299-315.
8. Kish SJ, Lerch J, Furukawa Y, Tong J, McCluskey T, et al. (2010) Decreased cerebral
cortical serotonin transporter binding in ecstasy users: a positron emission
tomography/[(11)C]DASB and structural brain imaging study. Brain 133: 1779-1797.
9. de Win MM, Jager G, Booij J, Reneman L, Schilt T, et al. (2008) Neurotoxic effects of
ecstasy on the thalamus. Br J Psychiatry 193: 289-296.

P á g i n a | 8 2
10. King GR, Ernst T, Deng W, Stenger A, Gonzales RM, et al. (2011) Altered brain
activation during visuomotor integration in chronic active cannabis users: relationship to
cortisol levels. J Neurosci 31: 17923-17931.
11. Pillay SS, Rogowska J, Kanayama G, Gruber S, Simpson N, et al. (2008) Cannabis and
motor function: fMRI changes following 28 days of discontinuation. Exp Clin
Psychopharmacol 16: 22-32.
12. Martín-Santos R, Fagundo AB, Crippa JA, Atakan Z, Bhattacharyya S, et al. (2010)
Neuroimaging in cannabis use: a systematic review of the literature. Psychol Med 40: 383-
98.
13. Goldstein RZ, Alia-Klein N, Tomasi D, Carrillo JH, Maloney T, et al. (2009) Anterior
cingulate cortex hypoactivations to an emotionally salient task in cocaine addiction. Proc
Natl Acad Sci U S A 106: 9453-9458.
14. Zubieta JK, Heitzeg MM, Xu Y, Koeppe RA, Ni L, et al. (2005) Regional cerebral blood
flow responses to smoking in tobacco smokers after overnight abstinence. Am J Psychiatry
162: 567-577.
15. Verdejo-García AJ, López-Torrecillas F, Aguilar de Arcos F, Pérez-García M (2005)
Differential effects of MDMA, cocaine, and cannabis use severity on distinctive components
of the executive functions in polysubstance users: a multiple regression analysis. Addict
Behav 30: 89-101.
16. Boellaard R, O’Doherty MJ, Weber WA, Mottaghy FM, Lonsdale MN, et al. (2010) FDG
PET and PET/CT: EANM procedure guidelines for tumour PET imaging: version 1.0. Eur J
Nucl Med Mol Imaging 37: 181-200.

P á g i n a | 8 3
17. Fehr C, Hohmann N, Gründer G, Dielentheis TF, Buchholz HG, et al. (2007) Tiagabine
does not attenuate alcohol-induced activation of the human reward system.
Psychopharmacology (Berl) 191: 975-983.
18. Volkow ND, Fowler JS, Wang GJ, Telang F, Logan J, et al. (2010) Cognitive control of
drug craving inhibits brain reward regions in cocaine abusers. Neuroimage 49: 2536-
2543.
19. Jovanovski D, Erb S, Zakzanis KK (2005) Neurocognitive deficits in cocaine users: a
quantitative review of the evidence. J Clin Exp Neuropsychol 27: 189-204.
20. Zakzanis KK, Campbell Z, Jovanovski D (2007) The neuropsychology of ecstasy
(MDMA) use: a quantitative review. Hum Psychopharmacol 22: 427-435.
21. Hanson KL, Cummins K, Tapert SF, Brown SA (2011) Changes in neuropsychological
functioning over 10 years following adolescent substance abuse treatment. Psychol Addict
Behav 25: 127-142
22. de Sola S, Tarancón T, Peña-Casanova J, Espadaler JM, Langohr K, et al. (2008) Auditory
event-related potentials (P3) and cognitive performance in recreational ecstasy polydrug
users: evidence from a 12-month longitudinal study. Psychopharmacology (Berl) 200:
425-437
23. Yuan Y, Zhu Z, Shi J, Zou Z, Yuan F, et al. (2009) Gray matter density negatively
correlates with duration of heroin use in young lifetime heroin-dependent individuals.
Brain Cogn 71: 223-228.
24. Breiter HC, Rosen BR (1999) Functional magnetic resonance imaging of brain reward
circuitry in the human. Ann N Y Acad Sci 877: 523-547.

P á g i n a | 8 4
25. Barrós-Loscertales A, Bustamante JC, Ventura-Campos N, Llopis JJ, Parcet MA, et al.
(2011) Lower activation in the right frontoparietal network during a counting Stroop task
in a cocaine-dependent group. Psychiatry Res 194: 111-118.
26. Bustamante JC, Barrós-Loscertales A, Ventura-Campos N, Sanjuán A, Llopis JJ, et al.
(2011) Right parietal hypoactivation in a cocaine-dependent group during a verbal
working memory task. Brain Res 1375: 111-119.
27. van Holst RJ, de Ruiter MB, van den Brink W, Veltman DJ, Goudriaan AE (2012) A
voxel-based morphometry study comparing problem gamblers, alcohol abusers, and
healthy controls. Drug Alcohol Depend. In press.
28. Sullivan EV, Deshmukh A, De Rosa E, Rosenbloom MJ, Pfefferbaum A (2005) Striatal
and forebrain nuclei volumes: contribution to motor function and working memory
deficits in alcoholism. Biol Psychiatry 57: 768-776.
29. Johnson-Greene D, Adams KM, Gilman S, Junck L (2002) Relationship between
neuropsychological and emotional functioning in severe chronic alcoholism. Clin
Neuropsychol 16: 300-309.
30. Zinn S, Stein R, Swartzwelder HS (2004) Executive functioning early in abstinence from
alcohol. Alcohol Clin Exp Res 28: 1338-1346.
31. Pitel AL, Perruchet P, Vabret F, Desgranges B, Eustache F, et al. (2009) The advantage
of errorless learning for the acquisition of new concepts' labels in alcoholics. Psychol Med
40: 497-502.
Tzourio-Mazoyer N, Landeau B, Papathanassiou D, Crivello F, Etard O, et al. (2002)
Automated anatomical labeling of activations in SPM using a macroscopic anatomical
parcellation of the MNI MRI single-subject brain. Neuroimage 15: 273-289.

P á g i n a | 8 5
* ARTÍCULO 3
Neural correlates of hot and cold executive functions in polysubstance addiction:
Association between neuropsychological performance and resting-PET brain
metabolism
* Moreno-López, L., Catena, A., Fernández-Serrano, M. J., Delgado-Rico, E., Stamatakis, E. A.,
Pérez-García, M., & Verdejo-García, A. (2012). Neural correlates of hot and cold executive
functions in polysubstance addiction: Association between neuropsychological
performance and resting-PET brain metabolism. Psychiatry Research: Neuroimaging, en
prensa.

P á g i n a | 8 6
1. Introduction
Contemporary neurobiological models consider addiction to be a brain disorder
that involves long-term neuroadaptations leading to persistent drug use despite
increasing negative consequences (Baler and Volkow, 2006; Goldstein and Volkow, 2002;
Verdejo-García et al., 2004). These neuroadaptations are thought to affect neural systems
involved in motivation, emotion, learning, memory and executive functioning (Ersche et
al., 2008; Verdejo-García and Bechara, 2009). From this perspective, it is crucial to
understand both the long-term neuropsychological consequences of drug abuse and their
neural substrates.
There is compelling evidence that substance dependent individuals (SDI) present
widespread deficits in neuropsychological functioning, and that these deficits are
especially prominent in so-called executive functions (Fernández-Serrano et al., 2011).
Executive functions are defined as a set of higher-order abilities involved in the
employment, monitoring, and regulation of goal-directed behaviors. Recent evidence
suggests that these functions can be classified into two broad domains: (i) cold executive
functions, involved in the processing of relatively abstract, context-free information, which
are linked to more dorsal and lateral regions of the prefrontal cortex, and (ii) hot executive
functions, which come to play when emotionally laden information is concerned, and are
linked to the functioning of the orbitofrontal cortex (OFC) (Kerr and Zelazo, 2004).
Although executive dysfunction in SDI has been extensively described, the study of
the neurobiological substrates of executive dysfunction in the context of drug addiction is
still a growing research area (Goldstein and Volkow, 2002; Lundqvist, 2010). A number of
functional magnetic resonance imaging (fMRI) studies have shown significant associations
between performance on cognitive tasks indexing cold aspects of executive functions and
abnormal activation of brain systems in SDI. Polysubstance abusers have shown
abnormally increased or decreased activation of the dorsolateral prefrontal cortex

P á g i n a | 8 7
(DLPFC), orbitofrontal cortex (OFC), temporo-parietal regions and the cerebellum during
performance on working memory (Desmond et al., 2003; Jager et al., 2006; Tomasi et al.,
2007), response inhibition (Bolla et al., 2004; Dao-Castellana et al., 1998; Eldreth et al.,
2004; Forman et al., 2004; Lee et al., 2005), and cognitive flexibility tasks (Gilman et al.,
1990; Goldstein et al., 2004). Nevertheless, very few studies have investigated the neural
correlates of indices of hot executive functions in SDI. These abilities have been associated
with the OFC and the cingulate cortex in cognitive neuroscience studies (Anderson et al.,
2006; Hartstra et al., 2010; Heberlein et al., 2008), and they have been found to be
dysfunctional in polysubstance abusers even after mid-term abstinence (circa six months)
(Fernández-Serrano et al., 2011). For example, two neuropsychological studies have
revealed deficits in goal-driven self-regulation in psychostimulant users using the Revised
Strategy Application Test (R-SAT) (Halpern et al., 2004; Verdejo-García et al., 2007), but
no studies to date have investigated the neural substrates of self-regulation in addiction.
Another key factor for addiction initiation and maintenance is adaptive decision-making,
which is typically indexed using the Iowa Gambling Task (IGT) (Bechara et al., 1994).
Performance on this task has been associated with hyperperfusion in the anterior
cingulate cortex (ACC), middle and superior frontal gyrus and DLPFC in small samples of
shortly abstinent cocaine-dependent individuals (Adinoff et al., 2003; Tucker et al., 2004).
However, because of the key role of decision-making in mid and long-term drug relapse
(Passetti et al., 2008; Paulus et al., 2005), there is a need to extend these findings in larger
samples of mid-term abstinent SDI. Similarly, the neural correlates of emotional
processing and perception must be considered in light of their association with hot aspects
of executive functions in SDI (Verdejo-García et al., 2007), and their predictive role in drug
relapse (Lubman et al., 2009). Only two previous neuroimaging studies have focused on
this domain, and they have found decreased ACC and amygdala activations in alcohol users
exposed to facial emotional expressions (Marinkovic et al., 2009; Salloum et al., 2007).

P á g i n a | 8 8
In this study we aimed to investigate the neuropsychological performance of a
sample of SDI and healthy controls (HC) on tests tapping on cold (i.e., updating, inhibition
and flexibility) and hot (i.e., self-regulation, decision-making and emotion perception)
aspects of executive functions, and the association between the neuropsychological
performance of SDI on those tests and PET-indexed regional metabolism. We hypothesize
that neuropsychological performance will be poorer in SDI than HC and that it will be
associated with DLPFC, ACC and temporal regions for tests of cold executive functions, and
with OFC, ACC and limbic regions for tests of hot executive function.
2. Methods
2.1. Participants
Forty-nine SDI were recruited from an inpatient treatment program at the centre
“Proyecto Hombre” in Granada (Spain). The treatment has a mean duration of 6 months
and integrates key elements of cognitive behavioral psychology and systemic approaches
to provide complete social rehabilitation and integration. Selection criteria for participants
in this study were (1) meeting the DSM-IV criteria for substance dependence at the onset
of the treatment; (2) absence of documented comorbid mood or personality disorders, as
assessed by clinical reports; (3) absence of documented head injury or neurological
disorders, as assessed by clinical reports and PET images; (4) to be literate enough to
perform any literacy tests, as assessed by the TAP reading skills test (Del Ser et al., 1997)
and (5) to have a minimum abstinence duration of 15 days before testing. In fact, the SDI
included in the sample had overall much longer abstinence duration (M=32.94, SD=11.25
weeks), allowing us to rule out acute or short term effects of drug intake. Since virtually all
participants had regularly abused two or more substances, the analyses were conducted
on the whole group and results were interpreted in relation to polysubstance use. Urine
toxicology screening (one-step Syva rapid tests) for the different drugs used were
conducted routinely in the SDI, so we can rule out the use of these substances during the

P á g i n a | 8 9
entire period of abstinence. None of the participants were experiencing withdrawal
symptoms before or during neuropsychological/PET testing as assessed by routine
medical examination. Likewise, none of the participants were enrolled in opioid
substitution treatments with methadone or other pharmacological treatments during the
course of the neuropsychological/PET testing. A group of drug-free healthy individuals
was recruited as a comparison group for neuropsychological performance, but these
participants were not PET-scanned. HC were recruited through local advertisements and
snowball communication among adult people from the community. Selection criteria for
these group were: (1) absence of current or past substance abuse, excluding past cannabis
use and past or current smoking or social drinking (in cases of current social drinking,
exclusion criteria included acute use of alcohol during the last 24 h before evaluation), (2)
absence of documented major psychiatric disorders, (3) absence of documented head
injury or neurological disorder and (4) not being on any medication that could affect the
normal neuropsychological functioning. This group only had exposure to alcohol use,
having a regular mean use of 8.85 standard units per week (SD=20.7) and a mean duration
of use of 6.7 years (SD=7.3). The main sociodemographic and clinical features from the
samples are displayed in Table 1.
Table 1 Sociodemographic data and clinical features of the participants.
SDI (n=49) HC (n=30)
Variable N % N %
Gender Male 41 84 24 80 Female 8 16 6 20
Mean SD Mean SD
Age (years) 32.67* 8.03 26.4* 8.03 Education (years) 9.73* 2.34 11.63* 2.04
* p ≤ 0.001

P á g i n a | 9 0
2.2. Testing protocols and procedures
This study was approved by the Human Subjects Committee of the University of
Granada. All participants signed an informed consent form before participating in the
study. The testing protocol, including the neuropsychological assessment and the PET
scanning were administered into two sessions that lasted approximately 70 minutes each.
The control group only participated in the first session. During the first session,
participants completed the Interview for Research on Addictive Behaviors and performed
the neuropsychological tests protocol. During the second session (up to 7 days later), SDI
were scanned at the nuclear medicine service of the “Virgen de las Nieves” hospital.
2.3. Instruments
2.3.1. Patterns of drug use
The information about the severity of drug addiction was collected by the
Interview for Research on Addictive Behavior (IRAB; (López-Torrecillas et al., 2001) (see
supplementary material). This is a brief interview that collects information about the
quantity (average dose), frequency (consumption episodes by month), and duration (years
of duration) of the use of a series of drugs of abuse, including cocaine, heroin, and alcohol.
The main drug use information of the SDI group is displayed in Table 2.

P á g i n a | 9 1
Table 2 Amount and duration of drug use in the substance dependence individuals.
Quantity of use
Substance Averagea S.D. Maximum Minimum
Cocaine (46) 48.69 44.83 180 0 Cannabis (37) 148.57 190.64 750 0 Heroin (16) 9.12 20.63 120 0 MDMA (18) 13.41 24.31 120 0 Alcohol (45) 571.24 489.34 1800 0
Duration of use (years)
Substance Duration S.D. Maximum Minimum Age of onset
Cocaine 7.95 5.95 23 0 23.26 Cannabis 8.31 8.13 29 0 20.91 Heroin 1.76 3.69 17 0 24.75 MDMA 1.40 2.28 8 0 23.66 Alcohol 10.85 7.67 27 0 20.87 Abstinenceb 32.94 11.25 60 12
a Average dose (per episode) × Frequency (per month). A dose is units in alcohol, grams in cocaine and heroin, fags in cannabis and pills in MDMA. b Abstinence in weeks. The number in brackets is the number of substance dependence individuals that consumed the substance.
2.3.2. Neuropsychological tests
We administered a battery of tests designed to assess several domains related to
cold and hot executive functions, including working memory, response inhibition,
cognitive flexibility, self-regulation, decision-making and emotion processing.
The tests used were Letter Number Sequencing (LNS) (Wechsler Adult Intelligence
Scale - WAIS-III, Wechsler, 1997), Stroop (Golden, 1978), Category Test (DeFilippis, 2002),
Revised Strategy Application Test (R-SAT) (Levine et al., 2000), Iowa Gambling Task (IGT)
(Bechara et al., 1994) and Ekman Faces Test (EFT) (Young et al., 2002). All tests were
administered in a fixed order and according to standard instructions. The description of
the instruments is provided in the supplementary material.
Due to technical problems during data acquisition, we could not obtain the
performance indices for some of the tests in the SDI group. Therefore, the number of SDI

P á g i n a | 9 2
participants included in the analyses of LNS was 40, 39 for Stroop, 47 for Category Test, 49
for R-SAT and IGT, and 48 for EFT.
2.3.3. PET image acquisition
PET scans were acquired with the ECAT/931 scanner (Siemens CTI ECAT EXACT),
at the nuclear medicine facility of the “Virgen de las Nieves” hospital in Granada, (Spain).
For each individual, we obtained 20 minute emission scans 30 minutes after injection of
FDG resting in quiet conditions. The subjects were scanned with their eyes open and ears
unplugged in a dimly illuminated quiet room. Raw data were processed using an iterative
reconstruction (OSEM method: 10 iterations, 32 subsets). Images were reoriented in
transaxial, coronal and saggital planes. For the analyses reported here the reconstructed
images used had 47 axial slices each with an in-plane resolution of 2.57 x 2.57 and a slice
thickness of 3.38mm.
2.4. Data analysis
2.4.1. Preprocessing of PET images
The PET images were converted from DICOM to NIFTI format, were spatially
normalized to the SPM5 PET template (Wellcome Department of Cognitive Neurology,
London, UK) using linear (12 affine) and non-linear normalization. An average of all the
normalized images was obtained and this acted as a study specific PET template. In this
manner we constructed a template in the same modality, from the same scanner as the
images we wanted to normalize. The raw PET images were then spatially normalized with
linear affine and nonlinear parameters to this study-specific template. Visual inspection
revealed this optimized spatial normalization technique produced satisfactory spatial
normalization for the PET image of every volunteer. Finally, the images were smoothed
with an 8mm3 isotropic Gaussian filter.
2.4.2. ROI Analysis

P á g i n a | 9 3
Having achieved satisfactory spatial normalisation we used the Marsbar tool
(http://marsbar.sourceforge.net- Brett et al., 2002) to extract 90 regions of interest (ROIs)
in the entire brain. We used the AAL atlas as a reference for these ROIs (Tzourio-Mazoyer
et al., 2002).
2.4.3. Statistical Analyses
2.4.3.1. Behavioral Analysis
ANCOVA models were conducted to test neuropsychological performance
differences between the groups while controlling for age and education in SPSS 15.0 for
Windows (SPSS Inc., Chicago IL). Cohen´s d estimations of effect sizes were calculated
according to formulations proposed in (Zakzanis, 2001). The association between length
of abstinence and neuropsychological performance was explored using bivariate
correlations in SPSS 15.0.
2.4.3.2. ROI analysis
Relative glucose uptake on standardized uptake value (SUV) were used to obtain
the mean uptake values for each ROI. Next, these values were correlated with the indices
of neuropsychological performance. In a second analysis, those ROIs that showed
significant associations with neuropsychological indices were also correlated with
estimates of severity of drug use. In both cases, we used partial correlations and age as
confounding variable in SPSS 15.0. Results are showed at p≤0.01 and p≤0.05 respectively
(uncorrected for multiple comparisons).
2.4.3.3. Voxel based analysis
Linear regressions were used to identify clusters with a significant correlation
between uptake values and indices of neuropsychological performance using age as
confounding factor. Scans were corrected for differences in global activity by including

P á g i n a | 9 4
proportional scaling in the regression model. We report local maximum peaks that survive
a voxel threshold of p<0.001 (uncorrected for multiple comparisons, cluster size > 100
voxels). Although this threshold offers only modest protection again risk of type I error, it
has been chosen in multiples PET studies under similar assumptions (i.e. Brooks et al.,
2009; Yaouhi et al., 2009). The relationship between duration of abstinence and resting
glucose metabolism was explored by using linear regression models but no associations
survived at a threshold of p<0.001, and therefore it was not further considered. We did not
use treatment seeking as a variable in our analyses because all our volunteers were on
treatment, and more specifically on the same modality of treatment (i.e., therapeutic
community). Finally, we carried out a second analysis in which the mean uptake of the
regions we found to be associated with neuropsychological performance in whole brain
voxel-based analysis were correlated with estimates of drug use using age as confounding
variable in SPSS 15.0. Results are showed at p≤0.01 and p≤0.05 respectively (uncorrected
for multiple comparisons).
3. Results
3.1. Neuropsychological performance
Results showed that SDI had significantly poorer performance than HC on all of the
neuropsychological tests administered (Table 3). Calculation of effect sizes showed large
effect sizes (d>0.8) for LNS (working memory), Stroop (response inhibition), Category
Test (cognitive flexibility), IGT (decision-making) and EFT (emotion processing), and
medium effect sizes (d>0.5) for R-SAT (self-regulation) (Table 3). We only found a
significant negative correlation between length of abstinence and performance on LNS (r=-
0.303).
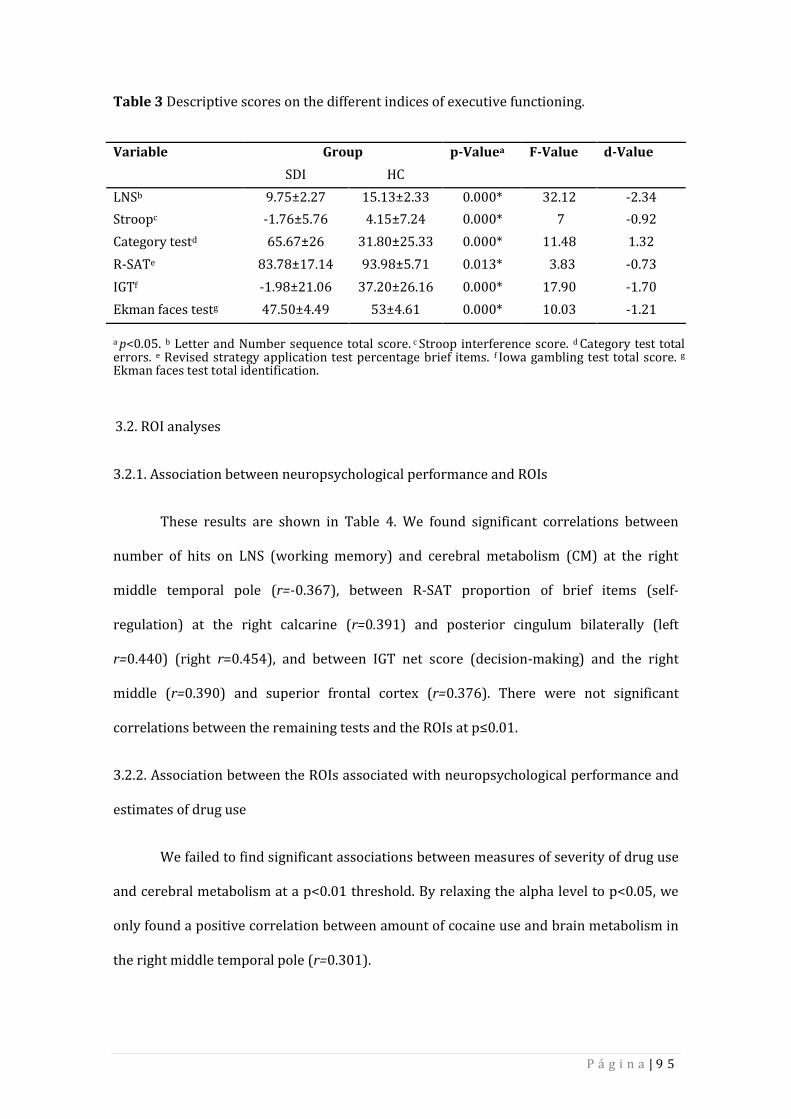
P á g i n a | 9 5
Table 3 Descriptive scores on the different indices of executive functioning.
Variable Group p-Valuea F-Value d-Value
SDI HC
LNSb 9.75±2.27 15.13±2.33 0.000* 32.12 -2.34
Stroopc -1.76±5.76 4.15±7.24 0.000* 7 -0.92
Category testd 65.67±26 31.80±25.33 0.000* 11.48 1.32
R-SATe 83.78±17.14 93.98±5.71 0.013* 3.83 -0.73
IGTf -1.98±21.06 37.20±26.16 0.000* 17.90 -1.70
Ekman faces testg 47.50±4.49 53±4.61 0.000* 10.03 -1.21
a p<0.05. b Letter and Number sequence total score. c Stroop interference score. d Category test total errors. e Revised strategy application test percentage brief items. f Iowa gambling test total score. g
Ekman faces test total identification.
3.2. ROI analyses
3.2.1. Association between neuropsychological performance and ROIs
These results are shown in Table 4. We found significant correlations between
number of hits on LNS (working memory) and cerebral metabolism (CM) at the right
middle temporal pole (r=-0.367), between R-SAT proportion of brief items (self-
regulation) at the right calcarine (r=0.391) and posterior cingulum bilaterally (left
r=0.440) (right r=0.454), and between IGT net score (decision-making) and the right
middle (r=0.390) and superior frontal cortex (r=0.376). There were not significant
correlations between the remaining tests and the ROIs at p≤0.01.
3.2.2. Association between the ROIs associated with neuropsychological performance and
estimates of drug use
We failed to find significant associations between measures of severity of drug use
and cerebral metabolism at a p<0.01 threshold. By relaxing the alpha level to p<0.05, we
only found a positive correlation between amount of cocaine use and brain metabolism in
the right middle temporal pole (r=0.301).

P á g i n a | 9 6
3.3. Voxel-based analyses
The main results from these analyses are summarized in Table 4. The results
presented in text and table correspond to an alpha level of p<0.001 (uncorrected, cluster
size > 100 voxels), but for demonstration purposes the figures display statistical
parametric maps thresholded at uncorrected voxel level of p<0.01.

P á g i n a | 9 7
Table 4 Relationship between neuropsychological performance and PET uptake using both ROI and voxel based methods.
L, left; R, right; (+) positive association; (-) negative association. Correlation ROI analyses are reported at p≤0.01 (uncorrected). Voxel-based analyses at p<0.001 (uncorrected, cluster size > 100 voxels). Talairach coordinates were obtained by applying the mni2tal function in MATLABR2006a (http://imaging.mrc-cbu.cam.ac.uk/imaging/MniTalairach) to the MNI coordinates obtained from SPM5. The corresponding anatomical names were obtained using the TalairachClient application – version 2.4.2. – (http://www.talairach.org/client.html) for peak and nearest grey matter.
Instrument ROIs Voxel-Based Analysis BA KE t p MNI Talairach
LNS R MiddleTemporal Pole (+) R Superior Temporal gyrus (+) BA22 621 5.23 0.000 52 -30 2 51 -29 3 R Inferior Temporal gyrus (+) BA37 261 3.77 0.000 52 -74 -4 51 -72 0 L Paracentral lobule (-) BA5 502 4.67 0.000 -20 -38 52 -20 -34 50 L Superior Frontal gyrus (-) BA8 449 3.95 0.000 -20 28 52 -20 30 46
Stroop
L Pallidus (-) 191 3.82 0.000 -16 -12 -4 -16 -12 -3 L Middle Frontal gyrus (-) BA10 160 3.72 0.000 -34 64 10 -34 62 6
Category test
L Superior Frontal gyrus (+) BA10 127 4.25 0.000 -26 68 4 -26 66 0 R Middle Frontal gyrus (-) BA6 268 3.59 0.000 50 10 48 50 12 44 L Inferior Parietal Lobule (-) BA40 198 3.74 0.000 -46 -48 38 -46 -45 37
R-SAT R Calcarine (+) R Posterior cingulate (+) L Posterior cingulate (+)
R Posterior cingulate (+) BA30 5811 4.12 0.000 18 -68 10 18 -65 12 R Anterior cingulate (+) BA32 702 3.75 0.000 14 32 16 14 32 13 R Precentral gyrus (+) BA6 416 3.96 0.000 22 -20 64 22 -16 60 R Inferior Temporal gyrus (+) BA20 152 3.58 0.000 40 2 -48 40 -0 -40 L Middle Temporal gyrus (-) BA21 464 4.48 0.000 -56 10 -20 -55 9 -17
IGT R Middle Frontal (+) R Superior Frontal (+)
R Middle Frontal gyrus (+) BA6 945 4.91 0.000 26 -6 60 26 -3 55 L Middle Frontal gyrus (+) BA6 377 3.71 0.000 -28 2 54 -28 4 50 R Middle Frontal gyrus (+) BA6 222 3.94 0.000 54 -12 56 53 14 51 L Superior Frontal gyrus (+) BA10 212 3.67 0.000 -18 68 14 -18 67 10 L Superior Frontal gyrus (+) BA8 141 3.72 0.000 -20 44 52 -20 45 46
Ekman Faces Test Lingual gyrus (-) BA18 308 4.10 0.000 0 -102 -14 0 -99 -7 Fusiform gyrus (-) BA19 191 3.90 0.000 -36 -78 -18 -36 -76 -11

P á g i n a | 9 8
3.3.1. Cold executive functions –Whole brain voxel-based correlates in SDI
For working memory, there was a positive correlation between LNS and one
cluster with peaks at the superior temporal gyrus (51 -29 3) and inferior temporal gyrus
(51 -72 0); this cluster included right superior and inferior lateral temporal regions
extending medially. LNS negatively correlated with a cluster in the left postcentral region
extending to parietal cortex, precuneus and paracentral lobule (peak at paracentral lobule,
-20 -34 50), and a second cluster which encompassed regions in the left frontal cortex
extending to the left DLPFC (peak at superior frontal gyrus, -20 30 46) (Figure 1 –Panel A).
For response inhibition, there was a negative correlation between the Stroop
interference score and two different clusters. The first cluster correspond to the left
thalamus, hippocampus and pallidum (peak at globus pallidus, -16 -12 -3), and the second
cluster encompassed the DLPFC bilaterally extending to the superior frontal cortex (peak
at middle frontal gyrus, -34 62 6) (Figure 1 –Panel B).
For cognitive flexibility, results showed a positive correlation between Category
Test total number of errors and a cluster which included the DLPFC and superior frontal
gyrus (peak at superior frontal gyrus, -26 66 0). There were also negative correlations
between task performance and two other clusters. The first was at the DLPFC extending to
the precentral gyrus with peak at middle frontal gyrus (50 12 44), and the second in the
parietal cortex extending to the supramarginal and angular area (peak at inferior parietal
lobule, -46 -45 37) (Figure 1 –Panel C).

P á g i n a | 9 9
Figure 1 Images showing the areas where cold executive functions correlated with brain
metabolism.
A. Letter Number sequencing negative (blue) and positive (yellow) correlations.
L -20 52 R
B. Stroop negative correlations (blue).
L -34 -16 R
C. Category Test negative (blue) and positive (yellow) correlations.
L -45 -26 45 R
In summary, working memory and cognitive flexibility performance negatively
correlated with cerebral metabolism in the left DLPFC and left parietal regions, whereas
response inhibition performance negatively correlated with bilateral DLPFC and basal
ganglia. We found positive correlations between working memory performance and

P á g i n a | 1 0 0
cerebral metabolism in the temporal gyrus, and between cognitive flexibility and cerebral
metabolism in the superior frontal gyrus.
3.3.2. Hot executive functions –Whole brain voxel-based correlates in SDI
For the R-SAT (self-regulation) results showed significant positive correlations
with four different clusters. One of them had a peak at the posterior cingulate (18 -65 12),
including bilateral middle and posterior cingulate cortex and right precuneus. A second
cluster had a statistical peak at the anterior cingulate (14 32 13) extending to the insula. A
third cluster encompassed regions of the precentral gyrus extending to superior frontal
cortex and to the postcentral gyrus and supplementary motor area (peak at precentral
gyrus 22 -16 60). The fourth cluster had a statistical peak at the inferior temporal gyrus
(40 -0 -40) extending to the temporal pole and fusiform. We also found a negative
correlation between R-SAT performance and a cluster localized in the temporal cortex
which included the middle and superior temporal gyrus and the temporal pole (peak at
the middle temporal gyrus -55 9 -17) (Figure 2 –Panel A).
For decision-making, there was a positive correlation between IGT scores and
three clusters. The first cluster included the DLPFC extending to posterior areas (peak at
the right middle frontal gyrus 26 -3 55), a second cluster which included the DLPFC
bilaterally (peak at left middle frontal gyrus -28 4 50 and right middle frontal gyrus 53 14
51) and two clusters with peak at the superior frontal gyrus (-18 67 10; -20 45 46) (Figure
2 –Panel B).
For emotion processing, there was a negative correlation between EFT emotion
recognition hits and two main clusters. One including the lingual gyrus (peak at lingual
gyrus, 0 -99 -7) and a second cluster including the cerebellum and fusiform and lingual
gyrus (peak at the fusiform gyrus -36 -76 -11) (Figure 2 –Panel C).

P á g i n a | 1 0 1
Figure 2 Images showing the areas where hot executive functions correlated with brain
metabolism.
A. Revised Strategy Application Test negative (blue) and positive (yellow) correlations.
L -56 8 35 R
B. Iowa Gambling Task positive correlations (yellow).
L -26 8 35 R
C. Ekman Faces Test negative correlations (blue).
L -37 -32 R
In summary, we found positive correlations between self-regulation performance
and cerebral metabolism within different sections of the cingulate cortex, superior frontal
cortex and inferior temporal gyrus, and between decision-making performance and
DLPFC/superior frontal metabolism. Conversely, we found negative correlations between

P á g i n a | 1 0 2
self-regulation performance and cerebral metabolism in the superior temporal gyrus, and
between emotion recognition performance and cerebral metabolism in the fusiform,
cerebellum and lingual gyrus.
3.3.3. The relationship between voxel-based derived regions associated with
neuropsychological performance and estimates of drug use
We only found a significant positive correlation between amount of heroin used
and metabolism in the left middle temporal gyrus (r=0.377) at p<0.01. By relaxing the
alpha level to p<0.05, we found positive correlations between duration of cannabis
(r=0.330) and cocaine used (r=0.296) and the right precentral gyrus, and negative
correlations between the amount of cannabis used and the left middle frontal gyrus (r=-
0.370) and left inferior parietal lobe (r=-0.307), the amount of heroin used and the right
inferior temporal gyrus (r=-0.308), and the amount of alcohol used and the right middle
frontal gyrus (r=-0.297).
4. Discussion
This study investigated the association between the neuropsychological
performance of SDI on measures of cold and hot executive functions (as measured outside
the scanner) and PET indexed regional brain glucose metabolism. SDI were also compared
to non-drug using controls with respect to behavioral performance in these tasks.
We found that SDI had significantly poorer performance than HC individuals on all
of the neuropsychological tests administered. Likewise, the neuropsychological
performance in the SDI was just affected by the duration of abstinence in the LNS task,
allowing us to rule out the effects of this variable on all other tasks. On the other hand, in
accordance with our initial hypotheses, we found that both cold and hot executive tests
were significantly associated with common brain regions, including the DLPFC, mid-
superior frontal gyrus and superior, inferior and middle temporal gyrus. All these regions

P á g i n a | 1 0 3
are thought to play important roles in the executive control of goal-driven behavior
(Bechara and Van Der Linden, 2005), and in neuroadaptations related to addiction (Koob
and Volkow, 2010); therefore, our findings highlight the relevance of this set of regions for
a wide range of cognitive functions that are significantly impaired in SDI. In terms of
differential associations, cold executive tests mainly correlated with frontal, temporal and
parietal regions, whereas hot executive functions correlated with midline mid-superior
frontal and cingulate regions in both ROI and whole brain voxel-based analyses. These
results are in agreement with the notion that different executive processes rely in both
common and partially dissociable neural networks (Collette et al., 2006), and that different
neural circuits are relevant to various aspects of abnormal cognitive functioning in SDI
(Verdejo-García et al., 2004). .
Specifically, our results showed that verbal working memory (indexed by LNS) is
mainly associated with DLPFC and temporo-parietal regions (Figure 1 –Panel A), which is
consistent with previous findings suggesting these regions are the key neural substrates
for memory updating and manipulation (Collette and Van der Linden, 2002; D’Esposito,
2007). For response inhibition, results showed associations between Stroop performance
and DLPFC and basal ganglia metabolism (Figure 1 –Panel B). This finding is in agreement
with evidence from cognitive and neuroimaging studies showing that fronto-striatal
alterations related to inhibitory control are ubiquitous across different forms of addiction,
and are associated with difficulties in ceasing drug use (Feil et al., 2010). For cognitive
flexibility results, we showed associations between the number of errors and DLPFC and
parietal cortex resting metabolism (Figure 1 –Panel C), replicating previous findings using
this probe and other switching perseveration tests (e.g., the Wisconsin Card Sorting Test)
in alcoholics (Adams et al., 1993) and healthy subjects (Thomas et al., 2011).
With regard to hot executive functions, results showed that R-SAT performance
was associated with resting metabolism in the mid-posterior and ACC, insula and temporal

P á g i n a | 1 0 4
regions (Figure 2 –Panel A). Both insula and ACC are involved in task-level control and
self-regulation of behavior (Nelson et al., 2010; Posner et al., 2007), the key cognitive
processes needed to develop and implement goal-driven adjustment of performance (as
indexed by this task). ACC and insula dysfunctions have been associated with stress-
induced craving, poor inhibitory control, impaired insight and less motivation to change in
SDI (Goldstein et al., 2009; Li and Sinha, 2008; Romero et al., 2010; Sinha and Li, 2007);
this range of symptoms may relate to core deficits of self-regulation (i.e., difficulties to set
and adjust behaviour according to long-term goals) in addicted individuals. Furthermore,
both anterior and posterior cingulate regions have been previously associated with
abnormal performance on multi-tasking measures, specifically with deficits in task
learning and remembering, rule-breaking and task switching (Burgess et al., 2000).
According to previous studies using similar methodologies, IGT performance in
mid-term abstinent SDI is associated with resting metabolism in both OFC and DLPFC
(Adinoff et al., 2003; Krain et al., 2006; Tucker et al., 2004) (Figure 2 –Panel B). This
finding is indicative of the notion that both cold processes, such as working memory or
switching skills relying on DLPFC functioning, and hot processes, such as monitoring of
reward value, relying on OFC functioning play a role in decision-making performance in
SDI (Bolla et al., 2003). Finally, although we failed to find significant associations between
ventral fronto-striatal regions and emotional perception, we found robust associations
between scores in this task and two areas (lingual and fusiform gyrus) (Figure 2 –Panel C)
that have been consistently associated with face processing in numerous studies
(Dinkelacker et al., 2011; Kanwisher and Yovel, 2006). The association between emotion
processing and the ventromedial frontal stream could be better grasped by fMRI studies
monitoring brain activation during actual emotional arousal.
Finally, we found significant associations between patterns of drug use and the
regions associated with executive performance in both ROI and whole brain voxel-based

P á g i n a | 1 0 5
analysis. Although mostly exploratory, some of these effects are informative about the link
between the use of particular substances and the brain regions supporting different
executive functions. For example, higher metabolism in the middle temporal gyrus was
associated with both R-SAT performance and severity of heroin use, in agreement with
data from resting fMRI showing left middle temporal gyrus dysfunction in chronic heroin
users (Jiang et al., 2011). Higher metabolism in the right precentral gyrus was associated
with both severity of cocaine use and cognitive flexibility, such that we may speculatively
suggest a role of this region in cocaine-induced motor perseveration deficits (Ersche et al.,
2008). Lower metabolism in the superior frontal gyrus was associated with both severity
of alcohol use and performance on cold executive tasks and on decision-making; this result
is fitting with the findings of Goldstein et al. (2004) who found that severity of alcohol use
was the main predictor of a composite index of executive impairment, which was
predicted by regional metabolism in the anterior cingulate. Finally, lower metabolism in
the adjacent middle frontal gyrus was linked to severity of cannabis use and performance
on response inhibition and decision-making, fitting with evidence of impairment of these
functions in chronic cannabis users (Battisti et al., 2010; Verdejo-Garcia et al., 2007).
These associations are in agreement with the notion that amount and duration of drug use
impact brain functioning in key regions for executive control, supporting the relevance of
the correlations between executive performance and brain metabolism in this clinical
group of SDI.
Several possible limitations of our study should be considered and addressed by
future research. In the first place, the methodology used in this study does not allow us to
monitor brain activity as a direct response to neuropsychological challenges; instead it
shows deficits in resting glucose metabolism that were related to test performance.
Therefore, this study cannot attribute NP function to an inability of the substance
dependent group to activate specific regions of the brain. FMRI or PET designs that involve
scanning during actual test performance may be better suited to test the neuronal

P á g i n a | 1 0 6
substrates of executive dysfunction in SDI. On the other hand, this design allowed us to
explore the brain correlates of highly complex neuropsychological paradigms, such as the
R-SAT that need to be simplified to fit them in fMRI designs. Furthermore, PET was
altogether not obtained in the control group and therefore it is not clear whether
performance differences on the NP tasks were associated with actual abnormalities (i.e.
compared to health) in glucose metabolism at rest. Future studies should examine if there
are differences in these associations between a clinical group and a control group. A fourth
limitation was the lack of estimates of nicotine use/dependence, considering that previous
studies have observed significant alterations in PET-indexed cerebral metabolism
associated with nicotine use (i.e. Rose et al., 2003; Zubieta et al., 2005) and should be
assess in any PET study of drug use. The voxel based analyses proved to be overall more
sensitive than the ROI method, this is an expected outcome because the voxel based
method is used to carry out statistical tests in smaller brain units (voxels) than the ROI
analysis (whole gyri). Finally, due to its correlational design, this study cannot yield
conclusions about cause-effect relationships in the area of the neuropsychology of drug
addiction.
Taken together, our results show that SDI have neuropsychological deficits in both
cold and hot executive functions. These deficits are associated with brain metabolism in
both common and partially dissociable neural networks encompassing frontal, parietal,
temporal and basal ganglia regions.

P á g i n a | 1 0 7
References
Adams, K.M., Gilman, S., Koeppe, R.A., Kluin, K.J., Brunberg, J.A., Dede, D., Berent, S., Kroll,
P.D., 1993. Neuropsychological deficits are correlated with frontal hypometabolism in
positron emission tomography studies of older alcoholic patients. Alcoholism, Clinical
and Experimental Research 17, 205-210.
Adinoff, B., Devous, M.D., Cooper, D.B., Best, S.E., Chandler, P., Harris, T., Cervin, C.A.,
Cullum, C.M., 2003. Resting regional cerebral blood flow and gambling task
performance in cocaine-dependent subjects and healthy comparison subjects. The
American Journal of Psychiatry 160, 1892-1894.
Anderson, S.W., Barrash, J., Bechara, A., Tranel, D., 2006. Impairments of emotion and real-
world complex behavior following childhood- or adult-onset damage to ventromedial
prefrontal cortex. Journal of the International Neuropsychological Society 12, 224-235.
Baler, R.D., Volkow, N.D., 2006. Drug addiction: the neurobiology of disrupted self-control.
Trends in Molecular Medicine 12, 559-566.
Battisti, R.A., Roodenrys, S., Johnstone, S.J., Pesa, N., Hermens, D.F., Solowij, N., 2010.
Chronic cannabis users show altered neurophysiological functioning on Stroop task
conflict resolution. Psychopharmacology (Berl) 212, 613-624.
Bechara, A., Damasio, A.R., Damasio, H., Anderson, S.W., 1994. Insensitivity to future
consequences following damage to the human prefrontal cortex. Cognition 50, 7-15.
Bechara, A., Van Der Linden, M., 2005. Decision-making and impulse control after frontal
lobe injuries. Current Opinion in Neurology 18, 734-739.
Bolla, K., Ernst, M., Kiehl, K., Mouratidis, M., Eldreth, D., Contoreggi, C., Matochik, J., Kurian,
V., Cadet, J., Kimes, A., Funderburk, F., London, E., 2004. Prefrontal cortical dysfunction

P á g i n a | 1 0 8
in abstinent cocaine abusers. The Journal of Neuropsychiatry and Clinical
Neurosciences 16, 456-464.
Bolla, K.I., Eldreth, D.A., London, E.D., Kiehl, K.A., Mouratidis, M., Contoreggi, C., Matochik,
J.A., Kurian, V., Cadet, J.L., Kimes, A.S., Funderburk, F.R., Ernst, M., 2003. Orbitofrontal
cortex dysfunction in abstinent cocaine abusers performing a decision-making task.
NeuroImage 19, 1085-1094.
Brett, M., Anton, J.L., Valabregue, R., Poline, J.B., 2002. Region of interest analysis using an
SPM toolbox. NeuroImage 16, 2.
Brooks, J.O. 3rd, Rosen, A.C., Hoblyn, J.C., Woodard, S.A., Krasnykh, O., Ketter, T.A., 2009.
Resting prefrontal hypometabolism and paralimbic hypermetabolism related to verbal
recall deficits in euthymic older adults with bipolar disorder. The American Journal of
Geriatric Psychiatry 17, 1022-1029.
Burgess, P.W., Veitch, E., de Lacy Costello, A., Shallice, T., 2000. The cognitive and
neuroanatomical correlates of multitasking. Neuropsychologia 38, 848-863.
Collette, F., Hogge, M., Salmon, E., Van der Linden, M., 2006. Exploration of the neural
substrates of executive functioning by functional neuroimaging. Neuroscience 139,
209-221.
Collette, F., Van der Linden, M., 2002. Brain imaging of the central executive component of
working memory. Neuroscience and Biobehavioral Reviews 26, 105-125.
Dao-Castellana, M.H., Samson, Y., Legault, F., Martinot, J.L., Aubin, H.J., Crouzel, C., Feldman,
L., Barrucand, D., Rancurel, G., Féline, A., Syrota, A., 1998. Frontal dysfunction in
neurologically normal chronic alcoholic subjects: metabolic and neuropsychological
findings. Psychological Medicine 28, 1039-1048.

P á g i n a | 1 0 9
DeFilippis, N.A., 2002. The Category Test, Computer Version. 2nd ed. Odessa, Florida,
Psychological Assessment Resources.
Del Ser, T., García-Montalvo, J.I., Martínez Espinosa, S., Delgado Villapalos, C., Bermejo, F.,
1997. The estimation of premorbid intelligence in Spanish people with the "Word
Accentuation Test" and its application to the diagnosis of dementia. Brain & Cognition
33, 343-356.
Desmond, J.E., Chen, S.H., DeRosa, E., Pryor, M.R., Pfefferbaum, A., Sullivan, E.V., 2003.
Increased frontocerebellar activation in alcoholics during verbal working memory: an
fMRI study. NeuroImage 19, 1510-1520.
D'Esposito, M., 2007. From cognitive to neural models of working memory. Philosophical
Transactions of the Royal Society B: Biological Sciences 29, 761-772.
Dinkelacker, V., Grüter, M., Klaver, P., Grüter, T., Specht, K., Weis, S., Kennerknecht, I., Elger,
C.E., Fernandez, G., 2010. Congenital prosopagnosia: multistage anatomical and
functional deficits in face processing circuitry. Journal of Neurology 258, 770-782.
Eldreth, D.A., Matochik, J.A., Cadet, J.L., Bolla, K.I., 2004. Abnormal brain activity in
prefrontal brain regions in abstinent marijuana users. NeuroImage 23, 914-920.
Ersche, K.D., Roiser, J.P., Robbins, T.W., Sahakian, B.J., 2008. Chronic cocaine but not
chronic amphetamine use is associated with perseverative responding in humans.
Psychopharmacology 197, 421-431.
Feil, J., Sheppard, D., Fitzgerald, P.B., Yücel, M., Lubman, D.I., Bradshaw, J.L., 2010.
Addiction, compulsive drug seeking, and the role of frontostriatal mechanisms in
regulating inhibitory control. Neuroscience and Biobehaviorals Reviews 35, 248-275.

P á g i n a | 1 1 0
Fernández-Serrano, M.J., Pérez-García, M., Verdejo-García, A., 2011. What are the specific
vs. generalized effects of drugs of abuse on neuropsychological performance?.
Neuroscience and Biobehavioral Reviews 35, 377-406.
Forman, S.D., Dougherty, G.G., Casey, B.J., Siegle, G.J., Braver, T.S., Barch, D.S., Stenger, V.A.,
Wick-Hull, C., Pisarov, L.A., Lorensen, E., 2004. Opiate addicts lack error-dependent
activation of rostral anterior cingulate. Biological Psychiatry 55, 531-537.
Gilman, S., Adams, K., Koeppe, R.A., Berent, S., Kluin, K.J., Modell, J.G., Kroll, P., Brunberg,
J.A., 1990. Cerebellar and frontal hypometabolism in alcoholic cerebellar degeneration
studied with positron emission tomography. Annals of Neurology 28, 775-785.
Golden, C.J., 1978. Stroop Color and Word Tets. A manual for clinical and experimental
uses. Wood Dale, Illinois, Stoelting Co.
Goldstein, R.Z., Craig, A.D., Bechara, A., Garavan, H., Childress, A.R., Paulus, M.P., Volkow,
N.D., 2009. The neurocircuitry of impaired insight in drug addiction. Trends in
Cognitive Sciences 9, 372-380.
Goldstein, R.Z., Leskowjan, A.C., Hoff, A.L., Hitzemann, R., Bashan, F., Khalsa, S., Wang, G.J.,
Fowler, J.S., Volkow, N.D., 2004. Severity of neuropsychological impairment in cocaine
and alcohol addiction: association with metabolism in the prefrontal cortex.
Neuropsychologia 42, 1447-1458.
Goldstein, R.Z., Volkow, N.D., 2002. Drug addiction and its underlying neurobiological
basis: neuroimaging evidence for the involvement of the frontal cortex. The American
Journal of Psychiatry 159, 1642-1652.
Halpern, J.H., Pope, H.G. Jr., Sherwood, A.R., Barry, S., Hudson, J.I., Yurgelun-Todd, D., 2004.
Residual neuropsychological effects of illicit 3,4-methylenedioxymethamphetamine

P á g i n a | 1 1 1
(MDMA) in individuals with minimal exposure to other drugs. Drug and Alcohol
Dependence 75, 135-147.
Hartstra, E., Oldenburg, J.F., Van Leijenhorst, L., Rombouts, S.A., Crone, E.A., 2010. Brain
regions involved in the learning and application of reward rules in a two-deck
gambling task. Neuropsychologia 48, 1438-1446.
Heberlein, A.S., Padon, A.A., Gillihan, S.J., Farah, M.J., Fellows, L.K., 2008. Ventromedial
frontal lobe plays a critical role in facial emotion recognition. Journal of Cognitive
Neuroscience 20, 721-733.
Jager, G., Kahn, R.S., Van Den Brink, W., Van Ree, J.M., Ramsey, N.F., 2006. Long-term effects
of frequent cannabis use on working memory and attention: an fMRI study.
Psychopharmacology 185, 358-368.
Jiang, G.H., Qiu, Y.W., Zhang, X.L., Han, L.J., Lv, X.F., Li, L.M., Lin, C.L., Zhuo, F.Z., Hu, S.Y., Tian,
J.Z., 2011. Amplitude low-frequency oscillation abnormalities in the heroin users: a
resting state fMRI study. Neuroimage 57,149-154.
Kanwisher, N., Yovel, G., 2006. The fusiform face area: a cortical region specialized for the
perception of faces. Philosophical Transactions of the Royal Society of London. Series
B, Biological Sciences 361, 2109-2128.
Kerr, A., Zelazo, P.D., 2004. Development of "hot" executive function: the children's
gambling task. Brain and Cognition 55, 148-157.
Koob, G.F., Volkow, N.D., 2010. Neurocircuitry of addiction. Neuropsychopharmacology 35,
217-238.
Krain, A.L., Wilson, A.M., Arbuckle, R., Castellanos, F.X., Milham, M.P., 2006. Distinct neural
mechanisms of risk and ambiguity: a meta-analysis of decision-making. NeuroImage
32, 477-484.

P á g i n a | 1 1 2
Lee, T.M., Zhou, W.H., Luo, X.J., Yuen, K.S., Ruan, X.Z., Weng, X.C., 2005. Neural activity
associated with cognitive regulation in heroin users: A fMRI study. Neuroscience
Letters 382, 211-216.
Levine, B., Dawson, D., Boutet, I., Schwartz, M.L., Stuss, D.T., 2000. Assessment of strategic
self-regulation in traumatic brain injury: its relationship to injury severity and
psychosocial outcome. Neuropsychology 14, 491-500.
Li, C.S., Sinha, R., 2008. Inhibitory control and emotional stress regulation: neuroimaging
evidence for frontal-limbic dysfunction in psycho-stimulant addiction. Neuroscience
and Biobehavioral Reviews 32, 581-597.
López-Torrecillas, F., Godoy, J.F., Pérez-García, M., Godoy, D., Sánchez-Barrera, M., 2001.
Variables modulating stress and coping that discriminate drug consumers from low or
not drug consumers. Addictive Behaviors 25, 161-165.
Lubman, D.I., Yücel, M., Kettle, J.W., Scaffidi, A., Mackenzie, T., Simmons, J.G., Allen, N.B.,
2009. Responsiveness to drug cues and natural rewards in opiate addiction:
associations with later heroin use. Archives of General Psychiatry 66, 205-212.
Lundqvist, T., 2010. Imaging cognitive deficits in drug abuse. Current Topics in Behavioral
Neurosciences 3, 247-275.
Marinkovic, K., Oscar-Berman, M., Urban, T., O’Reilly, C.E., Howard, J.A., Sawyer, K., Harris,
G.J., 2009. Alcoholism and dampened temporal limbic activation to emotional faces.
Alcoholism, Clinical and Experimental Research 33, 1880-1892.
Nelson, S.M., Dosenbach, N.U., Cohen, A.L., Wheeler, M.E., Schlaggar, B.L., Petersen, S.E.,
2010. Role of the anterior insula in task-level control and focal attention. Brain
Structure and Function 214, 669-680.

P á g i n a | 1 1 3
Passetti, F., Clark, L., Mehta, M.A., Joyce, E., King, M., 2008. Neuropsychological predictors
of clinical outcome in opiate addiction. Drug and Alcohol Dependence 94, 82-91.
Paulus, M.P., Tapert, S.F., Schuckit, M.A., 2005. Neural activation patterns of
methamphetamine-dependent subjects during decision making predict relapse.
Archives of General Psychiatry 62, 761-768.
Posner, M.I., Rothbart, M.K., Sheese, B.E., Tang, Y., 2007. The anterior cingulate gyrus and
the mechanism of self-regulation. Cognitive, Affective and Behavioral Neuroscience 7,
391-395.
Romero, M.J., Asensio, S., Palau, C., Sanchez, A., Romero, F.J., 2010. Cocaine addiction:
diffusion tensor imaging study of the inferior frontal and anterior cingulate white
matter. Psychiatry Research 181, 57-63.
Rose, J.E., Behm, F.M., Westman, E.C., Mathew, R.J., London, E.D., Hawk, T.C., Turkington,
T.G., Coleman, R.E., 2003. PET studies of the influences of nicotine on neural systems in
cigarette smokers. American Journal of Psychiatry 160, 323-333.
Salloum, J.B., Ramchandani, V.A., Bodurka, J., Rawlings, R., Momenan, R., George, D.,
Hommer, D.W., 2007. Blunted rostral anterior cingulate response during a simplified
decoding task of negative emotional facial expressions in alcoholic patients.
Alcoholism, Clinical and Experimental Research 31, 1490-1504.
Sinha, R., Li, C.S., 2007. Imaging stress- and cue-induced drug and alcohol craving:
association with relapse and clinical implications. Drug and Alcohol Review, 26, 25-31.
Thomas, L.A., Hall, J.M., Skup, M., Jenkins, S.E., Pine, D.S., Leibenluft, E., 2011. A
developmental neuroimaging investigation of the change paradigm. Developmental
Science 14, 148-161.

P á g i n a | 1 1 4
Tomasi, D., Goldstein, R.Z., Telang, F., Maloney, T., Alia-Klein, N., Caparelli, E.C., Volkow,
N.D., 2007. Widespread disruption in brain activation patterns to a working memory
task during cocaine abstinence. Brain Research 1171, 83-92.
Tucker, K.A., Potenza, M.N., Beauvais, J.E., Browndyke, J.N., Gottschalk, P.C., Kosten, T.R.,
2004. Perfusion abnormalities and decision making in cocaine dependence. Biological
Psychiatry 56, 527-530.
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello, F., Etard, O., Delcroix, N.,
Mazoyer, B., Joliot, M., 2002. Automated anatomical labelling of activations in SPM
using a macroscopic anatomical parcellation of the MNI MRI single subject brain.
NeuroImage 15, 273-289.
Verdejo-García, A., Bechara, A., 2009. A somatic marker theory of addiction.
Neuropharmacology 56, 48-62.
Verdejo-Garcia, A., Benbrook, A., Funderburk, F., David, P., Cadet, J.L., Bolla, K.I., 2007. The
differential relationship between cocaine use and marijuana use on decision-making
performance over repeat testing with the Iowa Gambling Task. Drug and Alcohol
Dependence 90, 2-11.
Verdejo-García, A., López-Torrecillas, F., Orozco, C., Pérez-García, M., 2004. Clinical
implications and methodological challenges in the study of the neuropsychological
correlates of cannabis, stimulant and opioid abuse. Neuropsychology Review 14, 1-41.
Verdejo-García, A., Rivas-Pérez, C., Vilar-López, R., Pérez-García, M., 2007. Strategic self-
regulation, decision-making and emotion processing in poly-substance abusers in their
first year of abstinence. Drug and Alcohol Dependence 12, 139-146.
Wechsler, D., 1997. Wechsler adult intelligence scale. 3rd ed. San Antonio, Texas, The
Psycological Corporation.

P á g i n a | 1 1 5
Yaouhi, K., Bertran, F., Clochon, P., Mézenge, F., Denise, P., Foret, J., Eustache, F.,
Desgranges, B., 2009. A combined neuropsychological and brain imaging study of
obstructive sleep apnea. Journal of Sleep Research 18, 36-48.
Young, A.W., Perrett, D.I., Calder, A.J., Sprengelmeyer, R., Ekman, P., 2002. Facial
expressions of emotion: stimuli and tests (FEEST). Bury St. Edmunds, Thames Valley
Test Company.
Zakzanis, K.K., 2001. Statistics to tell the truth, the whole truth, and nothing but the truth:
formulae, illustrative numerical examples, and heuristic interpretation of effects size
analyses for neuropsychological researchers. Archives of Clinical Neuropsychology 16,
653-677.
Zubieta, J.K., Heitzeg, M.M., Xu, Y., Koeppe, R.A., Ni, L., Guthrie, S., Domino, E.F., 2005.
Regional cerebral blood flow responses to smoking in tobacco smokers after overnight
abstinence. American Journal of Psychiatry 162, 567-577.

P á g i n a | 1 1 6
Supplementary Material
This document included the description of the Interview for Research on Addictive
Behaviour and the neuropsychological tasks used in the study.
The Interview for Research on Addictive Behaviour (IRAB; López-Torrecillas et al.,
2001) was used to examine the severity of drug use. This test evaluates, by means of a
brief interview, the quantity (average dose), frequency (consumption episodes by month),
and duration (years of duration) of the use of a series of drugs of abuse, including cocaine,
heroin, and alcohol, which were the drugs of abuse that motivated treatment requests in
the substance dependent individuals (SDI) included in this sample. For every substance
the participant had actually used, the following information was requested: (1) the
average dose of each target drug taken in each episode of use (number of grams for
cocaine and heroin and number of units for alcohol, considering that 25 cl. of hard liquor
(e.g., scotch) equals 2 units, while 25 cl. of wine or beer equals 1 unit); (2) the frequency of
substance use per month (daily, between one and three times upon a week, once a week,
between one and three times upon a month, or once a month); and (3) the number of
years that elapsed since the onset of the use. From this information, we obtained two
indices for each of the drugs used: (1) Amount (average dose x frequency), an index of
monthly use of the drug; and (2) Duration of drug use, measuring number of years of
exposure to the drug.
The description of the different neuropsychological tests used in the study is
detailed below.
Working memory: Letter Number Sequencing (LNS) (Wechsler Adult Intelligence
Scale -WAIS-III, Wechsler, 1997). The participant is read a sequence in which letters and
numbers are combined, and he or she is asked to reproduce the sequence heard, first
placing the numbers in ascending order and then the letters in alphabetical order. The test
consists of a series of elements of increasing complexity (progressively higher working

P á g i n a | 1 1 7
memory demands). Each element includes three trials, and in each trial the sequence is
read at one letter or number per second. The test is stopped when the participant misses
three trials of the same element. The main dependent variable used on this test was the
total number of hits.
Response inhibition: Stroop (Golden, 1978). This test consists of three forms, each
containing 100 elements distributed in five columns of 20 elements each. The first form is
made up of the words “RED”, “GREEN”, and “BLUE” ordered randomly and printed in black
ink. In this condition the participant is asked to read aloud, as quickly as possible, the
words written on this page in a time set at 45 seconds. The second form consists of strings
of “XXXX” printed in red, blue, or green ink. In this condition, the participant is asked to
overtly name as quickly as possible the color of these elements with a time limit of 45
seconds. The third form introduces the condition of interference, and it consists of the
words from the first form printed in the colors of the second. In this condition, the subject
is asked to name the color of the ink the word is written in, ignoring the word, also in 45
seconds. The main dependent variable used in this test was the interference score.
Cognitive flexibility: Category Test (DeFilippis, 2002). A computerized version of
this test was administered. The task consists of 208 stimuli that have different types of
designs grouped in seven subtests. For all the stimuli included in the same subtest, there is
an underlying rule that determines the appropriateness of the responses throughout this
subtest. However, this rule changes in the next subtest, so that the performance on the test
depends on the ability of the participant to infer these rules, and modify them when they
are no longer valid. Test instructions are intentionally ambiguous: we explained to the
participants that different types of designs will consecutively show up on the screen, and
that each design was associated with one of the first four numbers: 1, 2, 3, or 4. The
participant must press, for each stimulus, the key she/he thinks is associated with that
design; then the computer provides auditory feedback about the correctness of the

P á g i n a | 1 1 8
response. Therefore, participants have to use feedback to update response strategies
according to the changing rules, making this test a solid index of cognitive flexibility. The
main index of performance on the test was the total number of errors across the seven
subtests.
Self-regulation: Revised Strategy Application Test (R-SAT) (Levine et al., 2000). This
task is an unstructured paper and pencil test of strategy usage that is sensitive to
disturbed self-regulation. The R-SAT consisted of three simple activities: figure tracing,
sentence copying and object numbering. Each activity was displayed in two different
stacks (A and B), each containing 120 items. The items differed in two dimensions: size
(large vs. small) and the amount of time required to complete them (brief vs. medium vs.
long). Participants were instructed to perform these three activities during a 10 minute
time frame, trying to do some work on each of the stacks. The main goal of the task was to
win as many points as possible, considering that large items scored 0 points and small
items scored 100 points each. Additionally, completion of some items (those printed on
pages containing hand-drawn faces) led to the loss of all the points previously achieved.
Points were used in the instructions to see if participants would respond accordingly, but
the dependent variable in this task is the number of items and not the points. Given the
limited time, the most efficient strategy on this test (which the participant must discover)
is to complete the brief items and exclude the lengthy items. This requires the inhibition of
a tendency to complete all the items in sequence, which is established on the early pages,
where all the items are brief. The main dependent variable from the R-SAT was the
proportion of brief items completed with regard to the total number of items attempted.
Decision-making: Iowa Gambling Task (IGT) (Bechara et al., 1994). This task has
been proposed to be sensitive to real-life decision-making deficits. We used a
computerized version of the test (Bechara et al., 2001) four decks labelled A’, B’, C’, and D’,
with 60 cards each are displayed on the computer screen. The participant has to choose

P á g i n a | 1 1 9
cards from these decks during 100 trials with the aim of winning as much money as
possible, or in case she/he cannot win, at least try not to lose. Every time the participant
chooses a card she/he receives a varying amount of money and positive feedback (a
smiling face); however, in some cards, after winning she/he can also lose a varying
amount of money, accompanied by a frowning face. Decks C and D are considered
advantageous, because they provide little reward but also minimal punishment; leading to
substantial winning over time. On the other hand, decks A and B are considered
disadvantageous, because they provide high reward but also disproportionate losses;
leading to bankruptcy over time. The participant can actually track the money she/he wins
or loose watching a green bar on top of the screen. Good performance on the IGT implies
that the participant selects on average more cards from decks C’ and D’ (advantageous
decks: lower immediate gain but smaller future loss) than from decks A’ and B’
(disadvantageous decks: high immediate gain but larger future loss). Therefore, the
primary dependent variable from this task was the difference between the number of
cards selected from the advantageous minus the number of cards selected from the
disadvantageous decks: [(C+D)-(A+B)].
Emotion Recognition: Ekman Faces Test (EFT). This test is a computer task
assessing recognition of facial emotional expressions. The task uses stimuli from the
standardized test: “Facial Expressions of Emotion: Stimuli and Test” (FEEST; Young et al.,
2002). A series of 60 stimuli featuring faces portraying basic emotions were presented and
participants were asked to identify the emotion displayed by each face. Faces depicted
expressions of anger, disgust, fear, happiness, sadness and surprise (6 emotions, 10 faces
each). The total number of correct identifications was the main dependent variable from
this task.

P á g i n a | 1 2 0

P á g i n a | 1 2 1
[DISCUSIÓN]

P á g i n a | 1 2 2
1. Discusión General
La revisión de la literatura realizada al inicio de nuestra investigación nos permitió
subrayar la importancia que ciertos rasgos de personalidad como la impulsividad tenían
en las diferentes fases del proceso adictivo y la delimitación de ciertas inconsistencias y
limitaciones presentes en muchos de los estudios llevados a cabo en individuos
drogodependientes. Estas limitaciones, entre las que cabe destacar la repercusión
conjunta de factores disposicionales (rasgos premórbidos) y deterioros inducidos por el
consumo de drogas en la neurobiología de la adicción, los efectos del policonsumo de
distintas sustancias y los patrones heterogéneos de severidad de consumo de estas
sustancias presentes en estas poblaciones, han dificultado en gran medida la
interpretación de los resultados obtenidos por estudios previos, así como el
establecimiento de conclusiones sólidas sobre la asociación entre el consumo de drogas y
el cerebro.
Teniendo en cuenta estos antecedentes, el Objetivo General de esta Tesis Doctoral
fue el de investigar la asociación entre las alteraciones cerebrales estructurales y
funcionales presentes en individuos policonsumidores de drogas y (i) las variables de
personalidad que incrementan la predisposición al consumo y la dependencia, (ii) las
estimaciones de cantidad y duración de consumo de diferentes drogas, y (iii) el
funcionamiento neuropsicológico de los procesos ejecutivos típicamente afectados por el
consumo de drogas.
Este Objetivo General fue articulado en tres Objetivos Específicos que dieron lugar
a tres trabajos de investigación empírica y que han sido presentados en apartados
anteriores. Estos objetivos fueron: (i) estudiar la asociación entre las alteraciones
estructurales presentadas por un grupo de pacientes policonsumidores de drogas en
situación de abstinencia con consumo principal de cocaína y un conjunto de medidas de
personalidad, (ii) estudiar la asociación de la severidad del consumo de drogas con el

P á g i n a | 1 2 3
metabolismo cerebral en reposo de un grupo de pacientes policonsumidores de drogas en
situación de abstinencia y (iii) estudiar la asociación del funcionamiento neuropsicológico
y el metabolismo cerebral en situación de reposo en este mismo grupo de
policonsumidores de drogas.
Respecto al primer objetivo, nuestros resultados mostraron un patrón de
interacción diferente entre el volumen de materia gris regional del grupo de consumidores
y el grupo de individuos no consumidores y dos de las dimensiones de impulsividad
evaluadas: la urgencia negativa y la falta de premeditación. Estos resultados fueron
interpretados como el resultado de un patrón disfuncional en el grupo de consumidores
asociado con la alteración de los sistemas frontales y subcorticales propios de esta
población. Asimismo, se pudo demostrar la existencia de reducciones significativas en el
volumen regional de materia gris y materia blanca cerebral en estos consumidores, la
presencia de mayores puntuaciones en cuatro de las cinco dimensiones de personalidad
evaluadas y la asociación de los patrones de severidad de consumo de cocaína con índices
globales de materia gris cerebral.
El estudio de la asociación de la severidad del consumo de drogas con el
metabolismo cerebral permitió demostrar la existencia de asociaciones significativas entre
la cantidad de cocaína, alcohol y cannabis consumida y la duración del consumo de
heroína, alcohol y MDMA y el metabolismo regional cerebral de diferentes regiones
cerebrales. En concreto, se encontró que todas las drogas consumidas (excepto la cocaína)
mostraban una correlación negativa con el metabolismo de la corteza prefrontal
dorsolateral y que la severidad del consumo de alcohol y cocaína correlacionaba con el
metabolismo de la corteza promotora y el putamen y la corteza parietal respectivamente.
Estas asociaciones fueron interpretadas como el resultado de la existencia de mecanismos
de acción comunes y específicos de las distintas drogas sobre un conjunto de sistemas
neuroquímicos y estructuras cerebrales.

P á g i n a | 1 2 4
Por último, el estudio de la asociación del funcionamiento neuropsicológico y el
metabolismo cerebral permitió demostrar la existencia de asociaciones significativas entre
el rendimiento neuropsicológico de estos pacientes en tareas de funcionamiento ejecutivo
“frías” y “calientes” y un conjunto de regiones cerebrales. En este sentido, se encontró una
asociación significativa entre el rendimiento en las tareas de funcionamiento ejecutivo
“frías” y el metabolismo cerebral de la corteza prefrontal dorsolateral, la parte medial del
giro frontal superior, las partes superior e inferior de la corteza temporal y la corteza
parietal, y entre el rendimiento en las tareas de funcionamiento ejecutivo “calientes” y la
corteza prefrontal dorsolateral, la parte medial del giro frontal superior, la parte anterior y
posterior del cíngulo, la corteza temporal y el giro fusiforme. Estos resultados fueron
interpretados como el resultado de la diferente implicación de los sistemas cerebrales en
el funcionamiento ejecutivo y sus diferentes componentes. Asimismo, se pudo demostrar
la asociación entre el metabolismo cerebral y diferentes patrones de consumo y la
existencia de alteraciones neuropsicológicas en todos los procesos neuropsicológicos
evaluados al comparar la ejecución de nuestros pacientes con la de un grupo de individuos
no consumidores.
De estos resultados se derivan una serie de conclusiones y recomendaciones. En
primer lugar, la realización de estos tres estudios nos ha permitido profundizar en el
estudio de los factores explicativos de la asociación entre el consumo de drogas y las
alteraciones cerebrales y de funcionamiento neuropsicológico que se observan en esta
población. Los resultados encontrados demuestran que tanto las variables de
personalidad, en concreto las dimensiones de falta de premeditación y urgencia negativa,
como las variables relacionadas con el consumo de drogas se asocian con las alteraciones
encontradas en esta población. Así, la presencia de ciertos rasgos de personalidad como la
impulsividad, pueden predisponer al individuo vulnerable al inicio del consumo de drogas,
promover la transición del consumo recreativo a la dependencia, impedir que la persona
pueda abandonar el consumo a pesar de intentarlo en repetidas ocasiones y por último

P á g i n a | 1 2 5
provocar la recaída del consumidor abstinente (Verdejo-García et al., 2008), pero además,
el consumo prolongado de estas sustancias puede traer consigo la alteración del
funcionamiento cerebral y la exacerbación de la impulsividad resultado de la alteración de
las áreas asociadas con el control de la conducta (Ersche et al., 2010a).
La consideración de la impulsividad como un factor de riesgo para iniciarse en el
consumo de drogas ha sido avalada por diferentes tipos de estudios. En este sentido, se
han llevado a cabo investigaciones con diferentes poblaciones caracterizadas por
presentar un alto riesgo de iniciarse en el consumo de drogas (p.e. adolescentes, pacientes
con trastorno por déficits de atención e hiperactividad (TDAH) y otros trastornos
externalizantes o hijos de padres que han sido consumidores) o en jugadores patológicos,
un tipo de “adicción conductual” que comparte con las adicciones por sustancias
mecanismos etiológicos y de vulnerabilidad pero que se diferencia de estas en que en este
tipo de adicción no se produce la administración de una sustancia que puede causar
efectos dañinos a nivel cerebral (Bechara, 2003; Potenza, 2001) y se ha demostrado que
estas poblaciones presentan de forma consistente altas puntuaciones de impulsividad y
alteraciones en los procesos de control inhibitorio (Verdejo-García et al., 2008).
Sin embargo, tanto la impulsividad como la presencia de alteraciones
neuropsicológicas pueden incrementar como consecuencia directa del consumo de drogas.
Y es que, son múltiples los estudios que demuestran la existencia de alteraciones
significativas en los procesos de control inhibitorio y toma de decisiones en consumidores
de diferentes tipos de drogas y la asociación de estas con los diferentes patrones de
severidad del consumo evaluados.
Por otra parte, observamos que las diferentes sustancias evaluadas afectan de
forma generalizada a una serie de circuitos que han sido consistentemente asociados con
las alteraciones encontradas en esta población pero que también muestran cierta
especificidad. En este sentido, los resultados de éste y otros estudios parecen indicar que

P á g i n a | 1 2 6
múltiples sustancias producen deterioros globales similares sobre diversos mecanismos
de control ejecutivo pero que además, determinadas sustancias pueden producir
alteraciones más severas en componentes ejecutivos específicos (Fernández-Serrano et al.,
2011).
Es importante subrayar el hecho de que tanto el rendimiento en todas las pruebas
neuropsicológicas evaluadas como la severidad del consumo de todas las drogas
estudiadas (excepto de la cocaína), fueron asociadas con el metabolismo cerebral del área
prefrontal dorsolateral. La implicación de esta estructura en nuestra población coincide
con la evidencia que indica la asociación de esta estructura con múltiples procesos
ejecutivos y su alteración en consumidores de psicoestimulantes (Goldstein et al., 2004;
Makris et al., 2008b), opiáceos (Gerra et al., 1998), alcohol (Dao-Castellana et al., 1998;
Makris et al., 2008a) o cannabis (Eldreth et al., 2004). Es importante destacar la asociación
encontrada entre la cantidad de cannabis consumida y el metabolismo cerebral en esta
misma estructura. Y es que, hasta ahora, la mayoría de los estudios llevados a cabo con
esta población no han encontrado alteraciones significativas asociadas con el consumo de
cannabis en consumidores abstinentes de esta sustancia. Por lo que, estos resultados
subrayan la importancia de llevar a cabo nuevos estudios en esta población haciendo uso
de herramientas de neuroimagen cerebral, ya que estas han mostrado una mayor
sensibilidad en la detección de alteraciones en el funcionamiento cerebral aún en ausencia
de alteraciones a nivel conductual (p.e. Barrós-Loscertales et al., 2011a).
Asimismo, encontramos una asociación generalizada entre la severidad del
consumo de heroína, alcohol y MDMA y el metabolismo cerebral de la corteza temporal, un
área que ha sido encontrada alterada en los estudios llevados a cabo en estas poblaciones
y que coincide con las alteraciones en memoria encontradas en estos pacientes
(Fernández-Serrano et al., 2011).

P á g i n a | 1 2 7
Por último, es importante subrayar que los individuos drogodependientes
mostraron alteraciones en todos los procesos neuropsicológicos evaluados. La evidencia
indica que los consumidores de drogas presentan importantes déficits a nivel
neuropsicológico y que estos déficits son especialmente pronunciados en las llamadas
funciones ejecutivas (Fernández-Serrano et al., 2011). En este sentido, se ha demostrado
que los pacientes en tratamiento por drogodependencias con alteraciones en estos
procesos presentan un peor pronóstico y mayores tasas de recaídas que aquellos que no
presentan dichas alteraciones (Aharonovich et al., 2006; Grohman & Fals-Stewart, 1994;
Streeter et al., 2008). Es más, mientras que un adecuado funcionamiento ejecutivo ha sido
asociado con mayores probabilidades de mantenerse abstinente (Passetti et al., 2008), la
presencia de estas alteraciones se ha asociado con un menor nivel de implicación y
participación en los programas de tratamiento y con la severidad de los problemas
presentados por estos individuos en diferentes ámbitos de su funcionamiento cotidiano
(Kornreich et al., 2002; Verdejo-García, Bechara, Recknor, & Pérez-García, 2006) y ha
demostrado su validez en la predicción de recaídas de consumidores de diversas
sustancias (Aharonovich, Nunes, & Hasin, 2003; Passetti et al., 2008; Streeter et al., 2008;
Turner et al., 2009; Verdejo-García et al., 2012). Y es que, en comparación con otras
variables, las variables neuropsicológicas se asocian irremediablemente con las
neuroadaptaciones cerebrales que definen la naturaleza crónica de la adicción (Paulus et
al., 2005) y que se asocian tanto con la vulnerabilidad previa como con el daño provocado
por el consumo de las sustancias.
En este sentido, el conocimiento de los procesos neuropsicológicos afectados y sus
correspondientes correlatos neuroanatómicos puede llevarnos a desarrollar estrategias
más eficaces de tratamiento y rehabilitación, estrategias dirigidas al tratamiento de los
efectos generales y específicos asociados con cada tipo de droga y con las alteraciones en
el funcionamiento ejecutivo que sabemos que han sido asociadas con las recaídas. Un
tratamiento que podría verse favorecido por el uso combinado de estrategias dirigidas

P á g i n a | 1 2 8
específicamente al tratamiento de las funciones neuropsicológicas afectadas y de
tratamientos farmacológicos que hayan mostrado su validez en la mejora de ciertas
alteraciones cognitivas (Brady, Gray, & Tolliver, 2011; Sofuoglu, 2010) y que sin duda
alguna se verá favorecido por el uso de herramientas de neuroimagen cerebral.
2. Conclusiones
De los resultados obtenidos se derivan las siguientes conclusiones:
1. Los individuos policonsumidores de drogas con consumo principal de cocaína presentan
reducciones significativas del volumen regional de materia gris y materia blanca cerebral
en regiones fronto-límbicas y fronto-estriadas relacionadas con sus niveles de
impulsividad.
2. La asociación entre la impulsividad y el volumen regional de materia gris cerebral
muestra un patrón diferencial entre individuos policonsumidores de drogas con consumo
principal de cocaína y controles no consumidores: los niveles de impulsividad presentan
correlaciones positivas con el volumen de regiones relacionadas con la valoración de
reforzadores, una asociación que es inversa en el caso de los controles.
3. La severidad del consumo de diferentes drogas de abuso se asocia de forma general con
el metabolismo regional de la corteza prefrontal dorsolateral y la corteza temporal en
individuos policonsumidores de drogas.
4. La severidad del consumo de cocaína se asocia específicamente con el metabolismo
cerebral de la corteza parietal, la del alcohol con la corteza prefrontal y el putamen y la del
MDMA con el área postcentral en individuos policonsumidores de drogas.
5. Existen correlaciones significativas entre el rendimiento neuropsicológico de los
individuos policonsumidores de drogas en pruebas de funcionamiento ejecutivo y el
metabolismo cerebral de diversas regiones frontales y subcorticales.

P á g i n a | 1 2 9
6. Los individuos policonsumidores de drogas presentan deterioros equivalentes en las
funciones ejecutivas “frías” y “calientes” pero el rendimiento en las funciones ejecutivas
“frías” correlaciona específicamente con reducciones del metabolismo regional en áreas
frontales, temporales y parietales, mientras que el rendimiento en las funciones ejecutivas
“calientes” se correlaciona con el metabolismo regional de áreas frontales, temporales,
cíngulo y giro fusiforme.
3. Perspectivas de futuro
De los resultados obtenidos se derivan importantes perspectivas de investigación futura,
de entre las que cabe destacar:
1. Profundizar en la determinación de la dirección de causalidad de las alteraciones del
funcionamiento ejecutivo en individuos drogodependientes. Se hace necesario el diseño de
estudios longitudinales y en poblaciones de alto riesgo de iniciarse en el consumo de
drogas (por ejemplo en adolescentes, hijos de padres que han sido consumidores y niños y
adolescentes con trastornos de conducta o trastorno por déficit de atención e
hiperactividad), así como estudios transversales que incluyan variables genéticas,
neuropsicológicas y de neuroimagen cerebral.
2. Estudiar los efectos a largo plazo del consumo de diferentes sustancias utilizando
pruebas de funcionamiento neuropsicológico y de neuroimagen cerebral en consumidores
“puros” de las sustancias. En este sentido, aunque en nuestro estudio intentamos controlar
el efecto del co-abuso de las diferentes sustancias, el efecto del policonsumo no puede ser
completamente descartado (Halpern et al., 2004).
3. Profundizar en la investigación de los posibles efectos de programas de rehabilitación
específicos para cada tipo de consumidor.

P á g i n a | 1 3 0
4. Desarrollar paradigmas de neuroimagen funcional sensibles a la detección de
alteraciones en el funcionamiento cerebral de consumidores de cannabis en situación de
abstinencia.
5. Evaluar las alteraciones en los procesos emocionales de percepción y experiencia
emocional en consumidores de cannabis en situación de abstinencia.
6. Estudiar la validez de diferentes técnicas de neuroimagen cerebral en la predicción de
distintos índices de funcionamiento clínico, así como los potenciales beneficios de la
aplicación de estrategias de intervención sobre la rehabilitación de los individuos
drogodependientes.
7. Desarrollar programas de tratamiento dirigidos al reestablecimiento o la compensación
de las funciones neuropsicológicas y emocionales encontradas alteradas en los individuos
drogodependientes.

P á g i n a | 1 3 1
[DOCTORADO EUROPEO]

P á g i n a | 1 3 2
1. Summary
The review of the literature conducted at the beginning of our investigation
allowed us to emphasize the potential impact that premorbid personality traits, such as
impulsivity, may have on different stages of the addictive process and its neurobehavioral
correlates. We also observed a number of inconsistencies and limitations in the way that
polysubstance use and duration of abstinence was handled in many of the studies
conducted in this population (poor control of co-abuse of other drugs and predominance
of studies in acute or shortly abstinent drug users). In this study we attempted to address
some of these limitations by studying the association between brain structure and
function and impulsivity traits, severity of use of different drugs, and executive control
associated dysfunction in polysubstance dependent individuals during protracted
abstinence.
The main aim of this Thesis was therefore to investigate the association between
brain structure and function in substance dependent individuals and (i) trait impulsivity,
as a main risk factor and clinical symptom associated with addiction, (ii) lifetime estimates
of amount and duration of drugs used and (iii) performance on tests of executive
functioning, which are thought to be associated with drug-induced neuroadaptations.
This aim was subsequently decomposed in three specific aims, which correspond
to the three studies collected in this Thesis. The first aim was to investigate the association
between volumetric measures of gray and white matter (measured with Magnetic
Resonance Imaging) and trait impulsivity and estimates of cocaine use in cocaine-
dependent individuals. Here we predicted that both greater impulsivity and higher and
longer exposure to cocaine use would correlate with less gray and white matter in
prefrontal and subcortical structures. Our results showed a different pattern of interaction
between regional gray matter volume of the cocaine-dependent individuals and the
control group and two of the dimensions of impulsivity assessed: negative urgency and

P á g i n a | 1 3 3
lack of premeditation. These results were interpreted as a result of a dysfunctional pattern
in the cocaine-dependent group associated with the disruption of frontal and subcortical
systems in this population. Also, we found significant reductions in the volume of regional
gray and white matter in the cocaine-dependent individuals, the presence of higher scores
on four of the five personality dimensions assessed and the association of patterns of
severity of cocaine use and overall rates of cerebral gray matter. The results of this study
can be found in the article entitled “Trait impulsivity and prefrontal gray matter
reductions in cocaine dependent individuals” published in Drug and Alcohol Dependence.
The second aim was to investigate the association between brain regional
metabolism (measured with FDG-PET) and lifetime estimates of amount and duration of
cocaine, heroin, alcohol, MDMA and cannabis use in a sample of polysubstance dependent
individuals. Here we predicted that exposure to all substances would be significantly
associated with overlapping reductions of metabolism in frontal, limbic, striatal and
cerebellar regions, whereas cocaine and heroin use would specifically correlate with
parietal and temporal cortices respectively. We found that whereas all drugs consumed
(except cocaine) showed a negative correlation with the metabolism of the dorsolateral
prefrontal cortex (DLPFC), the severity of alcohol, cocaine and MDMA correlated with
regional metabolism in premotor cortex and putamen, parietal cortex and postcentral
gyrus respectively. These associations were interpreted as a result of the existence of
common and specific mechanisms of action of the different drugs studied on the
neurotransmitter and functional brain systems involved in addiction. The results of this
study can be found in the article entitled “Neural Correlates of the Severity of Cocaine,
Heroin, Alcohol, MDMA and Cannabis Use in Polysubstance Abusers: A Resting-PET Brain
Metabolism Study” published in PloS One.
Finally, the third aim was to investigate the association between brain regional
metabolism (measured with FDG-PET) and performance measures of executive

P á g i n a | 1 3 4
functioning in a sample of polysubstance dependent individuals. Here we predicted that
executive performance would be poorer in polysubstance users, and that it would be
associated with lateral prefrontal and temporal regions for tests of “cold” executive
functions, and with orbitofrontal cortex and limbic regions for tests of “hot” executive
function. Regression models showed that substance dependent individuals performance in
cold executive test was associated with regional brain metabolism in the DLPFC, mid-
superior frontal gyrus, superior and inferior temporal gyrus and inferior parietal cortex,
whereas performance in hot executive functioning was associated with DLPFC, mid-
superior frontal gyrus, anterior and mid-posterior cingulate, and temporal and fusiform
gyrus. These results are in agreement with the notion that different executive processes
rely in both common and partially dissociable neural networks (F Collette et al., 2006),
and that different neural circuits relevant to various aspect of abnormal cognitive
functioning in substance dependent individuals (Verdejo-García et al., 2004). Likewise, we
found an association between cerebral metabolism and the different pattern of use
showed by the substance dependent individuals and the existence of neuropsychological
impairments in all the processes evaluated. Our results can be found in the article entitled
“Neural correlates of hot and cold executive functions in polysubstance addiction:
association between neuropsychological performance and resting-PET brain metabolism”
published in Psychiatry Research: Neuroimaging.
From these results we can derive a set of conclusions and recommendations. First
of all, the implementation of these three studies has provided further evidence of the
factors that contribute to explain the association between drug use and brain functioning.
The results found indicate that both impulsivity and drug use are associated with brain
alterations in this population. Therefore, some aspects of substance dependence and its
related neuroadaptations may be associated with personality traits. During substance
dependence, drug users may perseverate in drug-taking behaviours and the prolonged use

P á g i n a | 1 3 5
of these substances can lead to different neuroadaptations or to the exacerbation of the
ones originally associated with impulse control (Ersche et al., 2010a).
Moreover, we found significant general and specific associations of the severity of
drug use and the regional brain metabolism in the substance dependence individuals.
These associations can be interpreted as a result of the existence of common and specific
mechanisms of action of the drug studied on neurotransmitter and brain systems involved
in the addiction cycle. Studies exploring the effects of specific rehabilitation programs
based on the common and specific effects of each type of substance used can be very
useful to improve novel tailored interventions. Likewise, it is important to underscore that
both the deficient performance on the neuropsychological tests of executive function and
the severity of use of all drugs studied (except cocaine) were associated with the
functioning of the DLPFC. The relevance of this structure for substance dependence
overlaps with its relevance for the functioning of multiple executive processes. Therefore,
our results are in agreement with those of previous studies showing significant alterations
of this region in users of psychostimulants (Goldstein et al., 2004; Makris et al., 2008b),
opiates (Gerra et al., 1998), alcohol (Dao-Castellana et al., 1998; Makris et al., 2008a) or
cannabis (Eldreth et al., 2004). Importantly, the amount of cannabis used also correlated
with the regional brain metabolism in this structure, pointing to a link between use of this
substance and neuroadaptations in this region even after long-term abstinence. These
results underscore the importance of conducting further studies in cannabis users
employing brain imaging tools, as these have shown a greater sensitivity in detecting
alterations in brain functioning even in the absence of behavioral alterations (e.g.
Loscertales Barros et al., 2011).
Finally, it is important to note that substance dependent individuals showed
alterations in all the neuropsychological processes assessed. These results agree with
those of previous studies that had detected generalized alterations in the executive

P á g i n a | 1 3 6
functions in users of several substances (Di Sclafani, Tolou-Shams, Price, & Fein, 2002;
Goldstein et al., 2004; Noël et al., 2001). Furthermore, these results may have important
clinical implications for the treatment and rehabilitation of substance dependent
individuals. First, previous studies have shown that neuropsychological alterations of
executive functions are associated with reduced level of involvement and participation of
substance dependent individuals in treatment programs, and with a higher rate of
dropping out of these programs (Aharonovich et al., 2006; Passetti et al., 2008; Streeter et
al., 2008; Turner et al., 2009). On the other hand, these alterations are also related to the
severity of the problems showed by substance dependent individuals in diverse aspects of
daily functioning, including employment, social, family and legal problems (Kornreich et
al., 2002; Verdejo-García et al., 2006). Therefore, it is possible that the clinical and daily
functioning of the substance dependent individuals would benefit from the application of
specific neuropsychological training and rehabilitation strategies.
From the results obtained, we can derive the following conclusions and future
perspectives:
2. Conclusions
1. Cocaine dependent individuals have lowered gray and white matter volume in
fronto-limbic and fronto-striatal regions related to their levels of impulsivity.
2. The association between impulsivity and regional gray matter volumes shows a
differential pattern in cocaine dependent individuals vs. healthy controls:
impulsivity levels have a positive correlation with the volumes of regions
associated with reinforcement valuation, whereas this association is negative in
the healthy control group.
3. The severity of use of different drugs is associated with overlapping prefrontal and
temporal regions metabolic reductions in substance dependent individuals.

P á g i n a | 1 3 7
4. The severity of cocaine use is specifically negatively correlated with brain metabolism in
the parietal cortex, the severity of alcohol with the premotor cortex and the putamen, and
the severity of MDMA with the postcentral gyrus.
5. There are significant correlations between substance dependent individuals’
neuropsychological performance and the brain metabolism of several frontal and
subcortical regions.
6. Substance dependent individuals’ performance in cold executive test is associated with
regional brain metabolism in the DLPFC, mid-superior frontal gyrus, superior and inferior
temporal gyrus and inferior parietal cortex, whereas performance in hot executive
functioning is associated with DLPFC, mid-superior frontal gyrus, anterior and mid-
posterior cingulate, and temporal and fusiform gyrus.
3. Future perspectives
1. To examine the direction of causality of the link between executive deficits and drug
addiction. One possible approach to this issue is to perform longitudinal studies able to
examine possible executive alterations in populations at high risk of initiating drug use
(for example: adolescents, children and adolescents with ADHD or conduct disorder, etc.).
Another possible approach is to explore the relationship between mechanisms of genetic
vulnerability and neuropsychological alterations in substance dependent individuals.
2. To examine the specific brain systems involved in the abnormal functioning of executive
and emotional processes in “pure” substance dependent individuals. In this respect,
although in our study we try to control the effect of co-abuse of the different substances
used, the effect of polydrug use can not be completely ruled out (Halpern et al., 2004).
3. To investigate the effects of specific rehabilitation programs for each type of substance
dependent individual.

P á g i n a | 1 3 8
3. To develop neuroimaging paradigms sensitive to detect alterations in the brain
functioning of cannabis dependent individuals after prolonged abstinence.
4. To study the alteration of the emotional processing in cannabis dependent individuals
after prolonged abstinence.
5. To study the validity of different brain imaging techniques for the prediction of different
clinical outcomes and for prediction of relapse.
6. To develop treatment programs aimed at the restoration or compensation of the
neuropsychological functions found altered in these populations.

P á g i n a | 1 3 9
[REFERENCIAS]

P á g i n a | 1 4 0
Adams, K. M., Gilman, S., Koeppe, R. A., Kluin, K. J., Brunberg, J. A., Dede, D., Berent, S., et al.
(1993). Neuropsychological deficits are correlated with frontal hypometabolism in
positron emission tomography studies of older alcoholic patients. Alcoholism, clinical
and experimental research, 17, 205-210.
Adinoff, B., Devous, M. D., Cooper, D. B., Best, S. E., Chandler, P., Harris, T., Cervin, C. A., et al.
(2003). Resting regional cerebral blood flow and gambling task performance in
cocaine-dependent subjects and healthy comparison subjects. The American journal
of psychiatry, 160, 1892-1894.
Aguilar de Arcos, F., Verdejo-Garcia, A., Lopez Jimenez, A., Montañez Pareja, M., Gomez
Juarez, E., Arraez Sanchez, F., & Perez-Garcia, M. (2008). Cambios en la respuesta
emocional ante estímulos visuales de contenido sexual en adictos a drogas.
Adicciones, 20, 117-124.
Aguilar de Arcos, F., Verdejo-García, A., Peralta-Ramírez, M. I., Sánchez-Barrera, M., &
Pérez-García, M. (2005). Experience of emotions in substance abusers exposed to
images containing neutral, positive, and negative affective stimuli. Drug and alcohol
dependence, 78, 159-167.
Aharonovich, E., Amrhein, P. C., Bisaga, A., Nunes, E. V., & Hasin, D. S. (2008). Cognition,
Commitment Language, and Behavioral Change Among Cocaine-Dependent Patients.
Psychology of Addictive Behaviors, 22, 557-562.
Aharonovich, E., Hasin, D. S., Brooks, A. C., Liu, X., Bisaga, A., & Nunes, E. V. (2006).
Cognitive deficits predict low treatment retention in cocaine dependent patients.
Drug and alcohol dependence, 81, 313-322.
Aharonovich, E., Nunes, E., & Hasin, D. (2003). Cognitive impairment, retention and
abstinence among cocaine abusers in cognitive-behavioral treatment. Drug and
alcohol dependence, 71, 207-211.
Albein-Urios, N., Martinez-González, J. M., Lozano, O., Clark, L., & Verdejo-García, A. (2012).
Comparison of impulsivity and working memory in cocaine addiction and
pathological gambling: Implications for cocaine-induced neurotoxicity. Drug and
alcohol dependence, en prensa.

P á g i n a | 1 4 1
Alia-Klein, N., Parvaz, M. A., Woicik, P. A., Konova, A. B., Maloney, T., Shumay, E., Wang, R.,
et al. (2011). Gene x disease interaction on orbitofrontal gray matter in cocaine
addiction. Archives of general psychiatry, 68, 283-294.
Anderson, S. W., Barrash, J., Bechara, A., & Tranel, D. (2006). Impairments of emotion and
real-world complex behavior following childhood- or adult-onset damage to
ventromedial prefrontal cortex. Journal of the International Neuropsychological
Society, 12, 224-235.
Aron, A. R. (2007). The neural basis of inhibition in cognitive control. The Neuroscientist,
13, 214-228.
Aron, J. L., & Paulus, M. P. (2007). Location, location: using functional magnetic resonance
imaging to pinpoint brain differences relevant to stimulant use. Addiction, 102, 33-43.
Asensio, S., Romero, M. J., Palau, C., Sanchez, A., Senabre, I., Morales, J. L., Carcelen, R., et al.
(2010). Altered neural response of the appetitive emotional system in cocaine
addiction: an fMRI Study. Addiction biology, 15, 504-516.
Ashburner, J., & Friston, K. J. (2005). Unified segmentation. NeuroImage, 26, 839-851.
Brand, M., Rothbauer, M., Driessen, M., & Markowitsch, H. (2008). Executive functions and
risky decision-making in patients with opiate dependence. Drug and Alcohol
Dependence, 97, 64-72.
Baler, R. D., & Volkow, N. D. (2006). Drug addiction: the neurobiology of disrupted self-
control. Trends in molecular medicine, 12, 559-566.
Barratt, E. S. (1959). Anxiety and Impulsiveness Related to Psychomotor Efficiency.
Perceptual and Motor Skills, 9, 191-198.
Barrós-Loscertales, A., Bustamante, J. C., Ventura-Campos, N., Llopis, J. J., Parcet, M. A., &
Avila, C. (2011a). Lower activation in the right frontoparietal network during a
counting Stroop task in a cocaine-dependent group. Psychiatry research, 194, 111-
118.
Barrós-Loscertales, A., Garavan, H., Bustamante, J. C., Ventura-Campos, N., Llopis, J. J.,
Belloch, V., Parcet, M. A., et al. (2011b). Reduced striatal volume in cocaine-dependent
patients. NeuroImage, 56, 1021-1026.

P á g i n a | 1 4 2
Battisti, R. A., Roodenrys, S., Johnstone, S. J., Pesa, N., Hermens, D. F., & Solowij, N. (2010).
Chronic cannabis users show altered neurophysiological functioning on Stroop task
conflict resolution. Psychopharmacology, 212, 613-624.
Baumann, M. H., Wang, X., & Rothman, R. B. (2007). 3,4-Methylenedioxymethamphetamine
(MDMA) neurotoxicity in rats: a reappraisal of past and present findings.
Psychopharmacology, 189, 407-424.
Bechara, A, Damasio, A. R., Damasio, H., & Anderson, S. W. (1994). Insensitivity to future
consequences following damage to human prefrontal cortex. Cognition, 50, 7-15.
Bechara, A, Damasio, H., & Damasio, A. R. (2000). Emotion, decision making and the
orbitofrontal cortex. Cerebral cortex, 10, 295-307.
Bechara, A. (2003). Risky business: emotion, decision-making, and addiction. Journal of
gambling studies, 19, 23-51.
Bechara, A. (2005). Decision making, impulse control and loss of willpower to resist drugs:
a neurocognitive perspective. Nature neuroscience, 8, 1458-1463.
Bechara, A, Dolan, S., Denburg, N., Hindes, A., Anderson, S. W., & Nathan, P. E. (2001).
Decision-making deficits, linked to a dysfunctional ventromedial prefrontal cortex,
revealed in alcohol and stimulant abusers. Neuropsychologia, 39, 376-389.
Bechara, A., & Van Der Linden, M. (2005). Decision-making and impulse control after
frontal lobe injuries. Current opinion in neurology, 18, 734-739.
Belin, D., Mar, A. C., Dalley, J. W., Robbins, T. W., & Everitt, B. J. (2008). High impulsivity
predicts the switch to compulsive cocaine-taking. Science, 320, 1352-1355.
Bermpohl, F., Kahnt, T., Dalanay, U., Hägele, C., Sajonz, B., Wegner, T., Stoy, M., et al. (2010).
Altered representation of expected value in the orbitofrontal cortex in mania. Human
brain mapping, 31, 958-969.
Bickel, W. K., & Marsch, L. A. (2001). Toward a behavioral economic understanding of drug
dependence: delay discounting processes. Addiction, 96, 73-86.

P á g i n a | 1 4 3
Bjork, J. M., Hommer, D. W., Grant, S. J., & Danube, C. (2004). Impulsivity in abstinent
alcohol-dependent patients: relation to control subjects and type 1-/type 2-like traits.
Alcohol, 34, 133-150.
Blair, R. J. R. (2003). Facial expressions, their communicatory functions and neuro-
cognitive substrates. Philosophical transactions of the Royal Society of London. Series
B, Biological sciences, 358, 561-572.
Boellaard, R., O’Doherty, M. J., Weber, W. A., Mottaghy, F. M., Lonsdale, M. N., Stroobants, S.
G., Oyen, W. J. G., et al. (2010). FDG PET and PET/CT: EANM procedure guidelines for
tumour PET imaging: version 1.0. European journal of nuclear medicine and molecular
imaging, 37, 181-200.
Bolla, K. I., Eldreth, D. A., London, E. D., Kiehl, K. A., Mouratidis, M., Contoreggi, C., Matochik,
J. A., et al. (2003). Orbitofrontal cortex dysfunction in abstinent cocaine abusers
performing a decision-making task. NeuroImage, 19, 1085-1094.
Bolla, K., Ernst, M., Kiehl, K., Mouratidis, M., Eldreth, D., Contoreggi, C., Matochik, J., et al.
(2004). Prefrontal cortical dysfunction in abstinent cocaine abusers. The Journal of
neuropsychiatry and clinical neurosciences, 16, 456-464.
Bolla, K. I., Funderburk, F. R., & Cadet, J. L. (2000). Differential effects of cocaine and
cocaine alcohol on neurocognitive performance. Neurology, 54, 2285-2292.
Bolla, K. I., Eldreth, D. A., Matochik, J. A., & Cadet, J. L. (2005). Neural substrates of faulty
decision-making in abstinent marijuana users. NeuroImage, 26, 480-492.
Bowden-Jones, H., McPhillips, M., Rogers, R., Hutton, S., & Joyce, E. (2005). Risk-taking on
tests sensitive to ventromedial prefrontal cortex dysfunction predicts early relapse in
alcohol dependency: a pilot study. The Journal of neuropsychiatry and clinical
neurosciences, 17, 417-420.
Brady, K. T., Gray, K. M., & Tolliver, B. K. (2011). Cognitive enhancers in the treatment of
substance use disorders: clinical evidence. Pharmacology, biochemistry, and behavior,
99, 285-294.
Brand, M., Labudda, K., & Markowitsch, H. J. (2006). Neuropsychological correlates of
decision-making in ambiguous and risky situations. Neural networks, 19, 1266-1276.

P á g i n a | 1 4 4
Brett, M., Anton, J. L., Valabregue, R., & Poline, J. B. (2002). Region of interest analysis using
an SPM toolbox. NeuroImage, 16, 2.
Breiter, H. C., & Rosen, B. R. (1999). Functional magnetic resonance imaging of brain
reward circuitry in the human. Annals of the New York Academy of Sciences, 877, 523-
547.
Brooks, J. O., Rosen, A. C., Hoblyn, J. C., Woodard, S. A., Krasnykh, O., & Ketter, T. A. (2009).
Resting prefrontal hypometabolism and paralimbic hypermetabolism related to
verbal recall deficits in euthymic older adults with bipolar disorder. The American
journal of geriatric psychiatry, 17, 1022-1029.
Buchert, R., Obrocki, J., Thomasius, R., Väterlein, O., Petersen, K., Jenicke, L., Bohuslavizki,
K. H., et al. (2001). Long-term effects of “ecstasy” abuse on the human brain studied
by FDG PET. Nuclear medicine communications, 22, 889-897.
Buckholtz, J. W., Treadway, M. T., Cowan, R. L., Woodward, N. D., Li, R., Ansari, M. S.,
Baldwin, R. M., et al. (2010). Dopaminergic network differences in human impulsivity.
Science, 329, 532.
Burgess, P. W., Veitch, E., de Lacy Costello, A., & Shallice, T. (2000). The cognitive and
neuroanatomical correlates of multitasking. Neuropsychologia, 38, 848-863.
Bustamante, J. C., Barrós-Loscertales, A., Ventura-Campos, N., Sanjuán, A., Llopis, J. J.,
Parcet, M.-A., & Avila, C. (2011). Right parietal hypoactivation in a cocaine-dependent
group during a verbal working memory task. Brain research, 1375, 111-119.
Cairns, E. D., & Cammock, J. (2002). Test de emparejamiento de figuras conocidas: MFF-20.
Madrid: TEA Ediciones.
Calder, A. J., & Young, A. W. (2005). Understanding the recognition of facial identity and
facial expression. Nature reviews. Neuroscience, 6, 641-651.
Camí, J., & Farré, M. (2003). Drug addiction. The New England journal of medicine, 349, 975-
986.
Chanraud, S., Martelli, C., Delain, F., Kostogianni, N., Douaud, G., Aubin, H. J., Reynaud, M., et
al. (2007). Brain morphometry and cognitive performance in detoxified alcohol-

P á g i n a | 1 4 5
dependents with preserved psychosocial functioning. Neuropsychopharmacology, 32,
429-438.
Childress, A. R., Mozley, P. D., McElgin, W., Fitzgerald, J., Reivich, M., & O’Brien, C. P. (1999).
Limbic activation during cue-induced cocaine craving. The American journal of
psychiatry, 156, 11-18.
Childress, A. R., Ehrman, R. N., Wang, Z., Li, Y., Sciortino, N., Hakun, J., Jens, W., et al. (2008).
Prelude to passion: limbic activation by “unseen” drug and sexual cues. PloS one, 3,
e1506.
Clark, L., Robbins, T. W., Ersche, K. D., & Sahakian, B. J. (2006). Reflection impulsivity in
current and former substance users. Biological psychiatry, 60, 515-522.
Coffey, S. F., Gudleski, G. D., Saladin, M. E., & Brady, K. T. (2003). Impulsivity and rapid
discounting of delayed hypothetical rewards in cocaine-dependent individuals.
Experimental and clinical psychopharmacology, 11, 18-25.
Cohen, L. J., Gertmenian-King, E., Kunik, L., Weaver, C., London, E. D., & Galynker, I. (2005).
Personality measures in former heroin users receiving methadone or in protracted
abstinence from opiates. Acta psychiatrica Scandinavica, 112, 149-158.
Collette, F., Hogge, M., Salmon, E., & Van der Linden, M. (2006). Exploration of the neural
substrates of executive functioning by functional neuroimaging. Neuroscience, 139,
209-221.
Collette, F., & Van der Linden, M. (2002). Brain imaging of the central executive component
of working memory. Neuroscience and biobehavioral reviews, 26, 105-125.
Cowan, R. L., Lyoo, I. K., Sung, S. M., Ahn, K. H., Kim, M. J., Hwang, J., Haga, E., et al. (2003).
Reduced cortical gray matter density in human MDMA (Ecstasy) users: a voxel-based
morphometry study. Drug and alcohol dependence, 72, 225-235.
Crean, R. D., Crane, N. A., & Mason, B. J. (2011). An evidence based review of acute and
long-term effects of cannabis use on executive cognitive functions. Journal of
addiction medicine, 5, 1-8.
Cyders, M. A., Flory, K., Rainer, S., & Smith, G. T. (2009). The role of personality dispositions
to risky behavior in predicting first-year college drinking. Addiction, 104, 193-202.

P á g i n a | 1 4 6
Cyders, M. A., Smith, G. T., Spillane, N. S., Fischer, S., Annus, A. M., & Peterson, C. (2007).
Integration of impulsivity and positive mood to predict risky behavior: development
and validation of a measure of positive urgency. Psychological assessment, 19, 107-
118.
Dalley, J. W., Fryer, T. D., Brichard, L., Robinson, E. S. J., Theobald, D. E. H., Lääne, K., Peña,
Y., et al. (2007). Nucleus accumbens D2/3 receptors predict trait impulsivity and
cocaine reinforcement. Science, 315, 1267-1270.
Dao-Castellana, M. H., Samson, Y., Legault, F., Martinot, J. L., Aubin, H. J., Crouzel, C.,
Feldman, L., et al. (1998). Frontal dysfunction in neurologically normal chronic
alcoholic subjects: metabolic and neuropsychological findings. Psychological
medicine, 28, 1039-1048.
Davidson, R. J. (2002). Anxiety and affective style: Role of prefrontal cortex and amygdale.
Biological Psychiatry, 51, 68-80.
Davies, S. J. C., Pandit, S. A., Feeney, A., Stevenson, B. J., Kerwin, R. W., Nutt, D. J., Marshall, E.
J., et al. (2005). Is there cognitive impairment in clinically “healthy” abstinent alcohol
dependence? Alcohol and alcoholism, 40, 498-503.
Desmond, J. E., Chen, S. H. A., DeRosa, E., Pryor, M. R., Pfefferbaum, A., & Sullivan, E. V.
(2003). Increased frontocerebellar activation in alcoholics during verbal working
memory: an fMRI study. NeuroImage, 19, 1510-1520.
del Ser, T., García-Montalvo, J. I., Martínez Espinosa, S., Delgado Villapalos, C., & Bermejo, F.
(1997). The estimation of premorbid intelligence in Spanish people with the "Word
Accentuation Test" and its application to the diagnosis of dementia. Brain &
Cognition, 33, 343-356.
DeFilippis, N. A. (2002). Category Test: Computer Version Research Edition. Lutz, Florida:
Psychological Assessment Resources.
de Win, M. M. L., Jager, G., Booij, J., Reneman, L., Schilt, T., Lavini, C., Olabarriaga, S. D., et al.
(2008). Neurotoxic effects of ecstasy on the thalamus. The British journal of
psychiatry, 193, 289-296.
de Sola, S., Tarancón, T., Peña-Casanova, J., Espadaler, J. M., Langohr, K., Poudevida, S.,
Farré, M., et al. (2008). Auditory event-related potentials (P3) and cognitive

P á g i n a | 1 4 7
performance in recreational ecstasy polydrug users: evidence from a 12-month
longitudinal study. Psychopharmacology, 200, 425-437.
de Oliveira, L. G., Barroso, L. P., Silveira, C. M., Sanchez, Z. V. D. M., De Carvalho Ponce, J.,
Vaz, L. J., & Nappo, S. A. (2009). Neuropsychological assessment of current and past
crack cocaine users. Substance use & misuse, 44, 1941-1957.
Di Sclafani, V., Tolou-Shams, M., Price, L. J., & Fein, G. (2002). Neuropsychological
performance of individuals dependent on crack-cocaine, or crack-cocaine and
alcohol, at 6 weeks and 6 months of abstinence. Drug and alcohol dependence, 66,
161-171.
Dickman, S. J. (1990). Functional and dysfunctional impulsivity: personality and cognitive
correlates. Journal of personality and social psychology, 58, 95-102.
Dinkelacker, V., Grüter, M., Klaver, P., Grüter, T., Specht, K., Weis, S., Kennerknecht, I., et al.
(2011). Congenital prosopagnosia: multistage anatomical and functional deficits in
face processing circuitry. Journal of neurology, 258, 770-782.
Dom, G., De Wilde, B., Hulstijn, W., van den Brink, W., & Sabbe, B. (2006). Decision-making
deficits in alcohol-dependent patients with and without comorbid personality
disorder. Alcoholism, clinical and experimental research, 30, 1670-1677.
D’Esposito, M., Detre, J. A., Alsop, D. C., Shin, R. K., Atlas, S., & Grossman, M. (1995). The
neural basis of the central executive system of working memory. Nature, 378, 279-
281.
D’Esposito, M. (2007). From cognitive to neural models of working memory. Philosophical
transactions of the Royal Society of London. Series B, Biological sciences, 362, 761-772.
D’Souza, M. S., & Duvauchelle, C. L. (2008). Certain or uncertain cocaine expectations
influence accumbens dopamine responses to self-administered cocaine and non-
rewarded operant behavior. European neuropsychopharmacology, 18, 628-638.
Eckardt, M. J., Rohrbaugh, J. W., Rio, D. E., & Martin, P. R. (1990). Positron emission
tomography as a technique for studying the chronic effects of alcohol on the human
brain. Annals of medicine, 22, 341-345.

P á g i n a | 1 4 8
Eldreth, D. A., Matochik, J. A., Cadet, J. L., & Bolla, K. I. (2004). Abnormal brain activity in
prefrontal brain regions in abstinent marijuana users. NeuroImage, 23, 914-920.
Ersche, K. D., Barnes, A., Jones, P. S., Morein-Zamir, S., Robbins, T. W., & Bullmore, E. T.
(2011). Abnormal structure of frontostriatal brain systems is associated with aspects
of impulsivity and compulsivity in cocaine dependence. Brain, 134, 2013-2024.
Ersche, K. D., Bullmore, E. T., Craig, K. J., Shabbir, S. S., Abbott, S., Müller, U., Ooi, C., et al.
(2010b). Influence of compulsivity of drug abuse on dopaminergic modulation of
attentional bias in stimulant dependence. Archives of general psychiatry, 67, 632-644.
Ersche, K. D., Roiser, J. P., Robbins, T. W., & Sahakian, B. J. (2008). Chronic cocaine but not
chronic amphetamine use is associated with perseverative responding in humans.
Psychopharmacology, 197, 421-431.
Ersche, K. D., Turton, A. J., Pradhan, S., Bullmore, E. T., & Robbins, T. W. (2010a). Drug
addiction endophenotypes: impulsive versus sensation-seeking personality traits.
Biological psychiatry, 68, 770-773.
European Monitoring Centre for Drugs and Drug Addiction (2011). Annual report 2011:
The state of the drugs problem in Europe. Descargado de:
http://www.emcdda.europa.eu/publications/annual-report/2011 el 3 de Mayo de
2012.
Evenden, J. L. (1999). Varieties of impulsivity. Psychopharmacology, 146, 348-361.
Fehr, C., Hohmann, N., Gründer, G., Dielentheis, T. F., Buchholz, H. G., Chechko, N.,
Yakushev, I., et al. (2007). Tiagabine does not attenuate alcohol-induced activation of
the human reward system. Psychopharmacology, 191, 975-983.
Feil, J., Sheppard, D., Fitzgerald, P. B., Yücel, M., Lubman, D. I., & Bradshaw, J. L. (2010).
Addiction, compulsive drug seeking, and the role of frontostriatal mechanisms in
regulating inhibitory control. Neuroscience and biobehavioral reviews, 35, 248-275.
Fein, G., Klein, L., & Finn, P. (2004). Impairment on a simulated gambling task in long-term
abstinent alcoholics. Alcoholism, clinical and experimental research, 28, 1487-1491.

P á g i n a | 1 4 9
Fernández-Serrano, M. J., Perales, J. C., Moreno-López, L., Pérez-García, M., & Verdejo-
García, A. (2012). Neuropsychological profiling of impulsivity and compulsivity in
cocaine dependent individuals. Psychopharmacology, 219, 673-683.
Fernández-Serrano, M. J., Pérez-García, M., Perales, J. C., & Verdejo-García, A. (2010a).
Prevalence of executive dysfunction in cocaine, heroin and alcohol users enrolled in
therapeutic communities. European journal of pharmacology, 626, 104-112.
Fernández-Serrano, M. J., Pérez-García, M., Schmidt Río-Valle, J., & Verdejo-García, A.
(2010b). Neuropsychological consequences of alcohol and drug abuse on different
components of executive functions. Journal of psychopharmacology, 24, 1317-1332.
Fernández-Serrano, M. J., Pérez-García, M., & Verdejo-García, A. (2011). What are the
specific vs. generalized effects of drugs of abuse on neuropsychological performance?
Neuroscience and biobehavioral reviews, 35, 377-406.
Fernández-Serrano, M. J., Lozano, O., Pérez-García, M., & Verdejo-García, A. (2010c). Impact
of severity of drug use on discrete emotions recognition in polysubstance abusers.
Drug and alcohol dependence, 109, 57-64.
Fillmore, M. T., & Rush, C. R. (2002). Impaired inhibitory control of behavior in chronic
cocaine users. Drug and alcohol dependence, 66, 265-273.
Fishbein, D. H., Krupitsky, E., Flannery, B. A., Langevin, D. J., Bobashev, G., Verbitskaya, E.,
Augustine, C. B., et al. (2007). Neurocognitive characterizations of Russian heroin
addicts without a significant history of other drug use. Drug and alcohol dependence,
90, 25-38.
Fisk, J. E., & Sharp, C. A. (2004). Age-related impairment in executive functioning: updating,
inhibition, shifting, and access. Journal of clinical and experimental neuropsychology,
26, 874-890.
Foisy, M. L., Kornreich, C., Fobe, A., D’Hondt, L., Pelc, I., Hanak, C., Verbanck, P., et al. (2007).
Impaired emotional facial expression recognition in alcohol dependence: do these
deficits persist with midterm abstinence? Alcoholism, clinical and experimental
research, 31, 404-410.

P á g i n a | 1 5 0
Fontes, M. A., Bolla, K. I., Cunha, P. J., Almeida, P. P., Jungerman, F., Laranjeira, R. R.,
Bressan, R. A., et al. (2011). Cannabis use before age 15 and subsequent executive
functioning. The British journal of psychiatry, 198, 442-447.
Forman, S. D., Dougherty, G. G., Casey, B. J., Siegle, G. J., Braver, T. S., Barch, D. M., Stenger, V.
A., et al. (2004). Opiate addicts lack error-dependent activation of rostral anterior
cingulate. Biological psychiatry, 55, 531-537.
Fox, H. C., Jackson, E. D., & Sinha, R. (2009). Elevated cortisol and learning and memory
deficits in cocaine dependent individuals: relationship to relapse outcomes.
Psychoneuroendocrinology, 34, 1198-1207.
Franklin, T. R., Acton, P. D., Maldjian, J. A., Gray, J. D., Croft, J. R., Dackis, C. A., O’Brien, C. P.,
et al. (2002). Decreased gray matter concentration in the insular, orbitofrontal,
cingulate, and temporal cortices of cocaine patients. Biological psychiatry, 51, 134-
142.
Frigerio, E., Burt, D. M., Montagne, B., Murray, L. K., & Perrett, D. I. (2002). Facial affect
perception in alcoholics. Psychiatry research, 113, 161-171.
Galynker, I. I., Watras-Ganz, S., Miner, C., Rosenthal, R. N., Des Jarlais, D. C., Richman, B. L., &
London, E. (2000). Cerebral metabolism in opiate-dependent subjects: effects of
methadone maintenance. The Mount Sinai journal of medicine, 67, 381-387.
Galynker, I. I., Eisenberg, D., Matochik, J. A., Gertmenian-King, E., Cohen, L., Kimes, A. S.,
Contoreggi, C., et al. (2007). Cerebral metabolism and mood in remitted opiate
dependence. Drug and alcohol dependence, 90, 166-174.
Garavan, H., Pankiewicz, J., Bloom, A., Cho, J. K., Sperry, L., Ross, T. J., Salmeron, B. J., et al.
(2000). Cue-induced cocaine craving: neuroanatomical specificity for drug users and
drug stimuli. The American journal of psychiatry, 157, 1789-1798.
Gazdzinski, S., Durazzo, T. C., Studholme, C., Song, E., Banys, P., & Meyerhoff, D. J. (2005).
Quantitative brain MRI in alcohol dependence: preliminary evidence for effects of
concurrent chronic cigarette smoking on regional brain volumes. Alcoholism, clinical
and experimental research, 29, 1484-1495.
George, M. S., Anton, R. F., Bloomer, C., Teneback, C., Drobes, D. J., Lorberbaum, J. P., Nahas,
Z., et al. (2001). Activation of prefrontal cortex and anterior thalamus in alcoholic

P á g i n a | 1 5 1
subjects on exposure to alcohol-specific cues. Archives of general psychiatry, 58, 345-
352.
George, O., & Koob, G. F. (2010). Individual differences in prefrontal cortex function and
the transition from drug use to drug dependence. Neuroscience and biobehavioral
reviews, 35, 232-247.
Gerra, G., Calbiani, B., Zaimovic, A., Sartori, R., Ugolotti, G., Ippolito, L., Delsignore, R.,
Rustichelli, P., & Fontanesi, B. (1998). Regional cerebral blood flow and comorbid
diagnosis in abstinent opioid addicts. Psychiatry research, 83, 117-126.
Gerra, G., Zaimovic, A., Ferri, M., Zambelli, U., Timpano, M., Neri, E., Marzocchi, G. F., et al.
(2000). Long-lasting effects of (+/-)3,4-methylenedioxymethamphetamine (ecstasy)
on serotonin system function in humans. Biological psychiatry, 47, 127-136.
Giancola, P. R., & Tarter, R. E. (1999). Executive cognitive functioning and risk for
substance abuse. Psychological Science, 10, 203-205.
Gilman, S., Adams, K., Koeppe, R. A., Berent, S., Kluin, K. J., Modell, J. G., Kroll, P., et al.
(1990). Cerebellar and frontal hypometabolism in alcoholic cerebellar degeneration
studied with positron emission tomography. Annals of neurology, 28, 775-785.
Golden, C. J. (1978). Stroop color and Word tests: a manual for clinical and experimental
uses. Wood Dale, Illinois: Stoelting Co.
Goldstein, R. Z., Tomasi, D., Rajaram, S., Cottone, L. A., Zhang, L., Maloney, T., Telang, F., et
al. (2007). Role of the anterior cingulate and medial orbitofrontal cortex in
processing drug cues in cocaine addiction. Neuroscience, 144, 1153-1159.
Goldstein, R. Z., & Volkow, N. D. (2002). Drug addiction and its underlying neurobiological
basis: neuroimaging evidence for the involvement of the frontal cortex. The American
Journal of Psychiatry, 159, 1642-1652.
Goldstein, R. Z., & Volkow, N. D. (2011). Dysfunction of the prefrontal cortex in addiction:
neuroimaging findings and clinical implications. Nature reviews. Neuroscience, 12,
652-669.
Goldstein, R. Z., Alia-Klein, N., Tomasi, D., Carrillo, J. H., Maloney, T., Woicik, P. A., Wang, R.,
et al. (2009). Anterior cingulate cortex hypoactivations to an emotionally salient task

P á g i n a | 1 5 2
in cocaine addiction. Proceedings of the National Academy of Sciences of the United
States of America, 106, 9453-9458.
Goldstein, R. Z., Leskovjan, A. C., Hoff, A. L., Hitzemann, R., Bashan, F., Khalsa, S. S., Wang, G.-
J., et al. (2004). Severity of neuropsychological impairment in cocaine and alcohol
addiction: association with metabolism in the prefrontal cortex. Neuropsychologia,
42, 1447-1458.
Goudriaan, A. E., Oosterlaan, J., de Beurs, E., & van den Brink, W. (2006). Neurocognitive
functions in pathological gambling: a comparison with alcohol dependence, Tourette
syndrome and normal controls. Addiction, 101, 534-547.
Grant, D. A. & Berg, E. A. A. (1948). A behavioral analysis of degree of reinforcement and
ease of shifting to new responses in a Weigel-type card-sorting problem. Journal of
Experimental Psychology, 38, 404-411.
Grant, D. A., & Berg, E. A. (1993). Wisconsin card sorting test. California, Los Angeles:
Western Psychological Services.
Grant, I., Gonzalez, R., Carey, C. L., Natarajan, L., & Wolfson, T. (2003). Non-acute (residual)
neurocognitive effects of cannabis use: a meta-analytic study. Journal of the
International Neuropsychological Society, 9, 679-689.
Grant, S., Contoreggi, C., & London, E. D. (2000). Drug abusers show impaired performance
in a laboratory test of decision making. Neuropsychologia, 38, 1180-1187.
Green, A R, King, M. V., Shortall, S. E., & Fone, K. C. F. (2011). Lost in translation: preclinical
studies on MDMA provide information on mechanisms of action, but do not allow
accurate prediction of adverse events in humans. British journal of pharmacology, en
prensa.
Green, A. R., Mechan, A. O., Elliott, J. M., O’Shea, E., & Colado, M. I. (2003). The
pharmacology and clinical pharmacology of 3,4-methylenedioxymethamphetamine
(MDMA, “ecstasy”). Pharmacological reviews, 55, 463-508.
Gruber, S. A., Silveri, M. M., & Yurgelun-Todd, D. A. (2007). Neuropsychological
consequences of opiate use. Neuropsychology review, 17, 299-315.

P á g i n a | 1 5 3
Gruber, S. A., & Yurgelun-Todd, D. A. (2001). Neuropsychological correlatos of drug abuse.
In Kaufman, M.J. (Ed). Brain imaging in substance abuse: research, clinical and forensic
appliactions. Totowa: New Jersey: Human Press Inc.
Gu, H., Salmeron, B. J., Ross, T. J., Geng, X., Zhan, W., Stein, E. A., & Yang, Y. (2010).
Mesocorticolimbic circuits are impaired in chronic cocaine users as demonstrated by
resting-state functional connectivity. NeuroImage, 53, 593-601.
Halpern, J. H., Pope, H. G. J., Sherwood, A. R., Barry, S., Hudson, J. I., & Yurgelun-Todd, D.
(2004). Residual neuropsychological effects of illicit 3,4-
methylenedioxymethamphetamine (MDMA) in individuals with minimal exposure to
other drugs. Drug and alcohol dependence, 75, 135-147.
Hanson, K. L., Cummins, K., Tapert, S. F., & Brown, S. A. (2011). Changes in
neuropsychological functioning over 10 years following adolescent substance abuse
treatment. Psychology of addictive behaviors, 25, 127-142.
Hanson, K. L., Luciana, M., & Sullwold, K. (2008). Reward-related decision-making deficits
and elevated impulsivity among MDMA and other drug users. Drug and alcohol
dependence, 96, 99-110.
Hartstra, E., Oldenburg, J. F. E., Van Leijenhorst, L., Rombouts, S. A. R. B., & Crone, E. A.
(2010). Brain regions involved in the learning and application of reward rules in a
two-deck gambling task. Neuropsychologia, 48, 1438-1446.
Heberlein, A. S., Padon, A. A., Gillihan, S. J., Farah, M. J., & Fellows, L. K. (2008).
Ventromedial frontal lobe plays a critical role in facial emotion recognition. Journal of
cognitive neuroscience, 20, 721-733.
Heil, S. H., Johnson, M. W., Higgins, S. T., & Bickel, W. K. (2006). Delay discounting in
currently using and currently abstinent cocaine-dependent outpatients and non-
drug-using matched controls. Addictive behaviors, 31, 1290-1294.
Jang, D. P., Namkoong, K., Kim, J. J., Park, S., Kim, I. Y., Kim, S. I., Kim, Y. B., et al. (2007). The
relationship between brain morphometry and neuropsychological performance in
alcohol dependence. Neuroscience Letters, 428,21-26.

P á g i n a | 1 5 4
Jager, G., Kahn, R. S., Van Den Brink, W., Van Ree, J. M., & Ramsey, N. F. (2006). Long-term
effects of frequent cannabis use on working memory and attention: an fMRI study.
Psychopharmacology, 185, 358-368.
Jiang, G. H., Qiu, Y. W., Zhang, X. L., Han, L. J., Lv, X. F., Li, L. M., Lin, C. L., et al. (2011).
Amplitude low-frequency oscillation abnormalities in the heroin users: a resting state
fMRI study. NeuroImage, 57, 149-154.
Johnson-Greene, D., Adams, K. M., Gilman, S., & Junck, L. (2002). Relationship between
neuropsychological and emotional functioning in severe chronic alcoholism. The
Clinical neuropsychologist, 16, 300-309.
Jovanovski, D., Erb, S., & Zakzanis, K. K. (2005). Neurocognitive deficits in cocaine users: a
quantitative review of the evidence. Journal of clinical and experimental
neuropsychology, 27, 189-204.
Kalechstein, A. D., De La Garza, R., Mahoney, J. J., Fantegrossi, W. E., & Newton, T. F. (2007).
MDMA use and neurocognition: a meta-analytic review. Psychopharmacology, 189,
531-537.
Kanwisher, N., & Yovel, G. (2006). The fusiform face area: a cortical region specialized for
the perception of faces. Philosophical transactions of the Royal Society of London.
Series B, Biological sciences, 361, 2109-2128.
Kerr, A., & Zelazo, P. D. (2004). Development of “hot” executive function: the children’s
gambling task. Brain and cognition, 55, 148-157.
Kilts, C. D., Gross, R. E., Ely, T. D., & Drexler, K. P. G. (2004). The neural correlates of cue-
induced craving in cocaine-dependent women. The American journal of psychiatry,
161, 233-241.
King, G. R., Ernst, T., Deng, W., Stenger, A., Gonzales, R. M. K., Nakama, H., & Chang, L.
(2011). Altered brain activation during visuomotor integration in chronic active
cannabis users: relationship to cortisol levels. The Journal of neuroscience, 31, 17923-
17931.
Kirby, K. N., Petry, N. M., & Bickel, W. K. (1999). Heroin addicts have higher discount rates
for delayed rewards than non-drug-using controls. Journal of experimental
psychology. General, 128, 78-87.

P á g i n a | 1 5 5
Kirby, K. N., & Petry, N. M. (2004). Heroin and cocaine abusers have higher discount rates
for delayed rewards than alcoholics or non-drug-using controls. Addiction, 99, 461-
471.
Kish, S. J., Lerch, J., Furukawa, Y., Tong, J., McCluskey, T., Wilkins, D., Houle, S., et al. (2010).
Decreased cerebral cortical serotonin transporter binding in ecstasy users: a positron
emission tomography/[(11)C]DASB and structural brain imaging study. Brain, 133,
1779-1797.
Koob, G. F., & Volkow, N. D. (2010). Neurocircuitry of addiction.
Neuropsychopharmacology, 35, 217-238.
Kornreich, C., Blairy, S., Philippot, P., Hess, U., Noël, X., Streel, E., Le Bon, O., et al. (2001).
Deficits in recognition of emotional facial expression are still present in alcoholics
after mid- to long-term abstinence. Journal of studies on alcohol, 62, 533-542.
Kornreich, C., Foisy, M. L., Philippot, P., Dan, B., Tecco, J., Noël, X., Hess, U., et al. (2003).
Impaired emotional facial expression recognition in alcoholics, opiate dependence
subjects, methadone maintained subjects and mixed alcohol-opiate antecedents
subjects compared with normal controls. Psychiatry research, 119, 251-260.
Kornreich, C., Philippot, P., Foisy, M. L., Blairy, S., Raynaud, E., Dan, B., Hess, U., et al. (2002).
Impaired emotional facial expression recognition is associated with interpersonal
problems in alcoholism. Alcohol and alcoholism, 37, 394-400.
Krain, A. L., Wilson, A. M., Arbuckle, R., Castellanos, F. X., & Milham, M. P. (2006). Distinct
neural mechanisms of risk and ambiguity: a meta-analysis of decision-making.
NeuroImage, 32, 477-484.
Kreek, M. J., Bart, G., Lilly, C., LaForge, K. S., & Nielsen, D. A. (2005). Pharmacogenetics and
human molecular genetics of opiate and cocaine addictions and their treatments.
Pharmacological reviews, 57, 1-26.
Lane, S. D., Steinberg, J. L., Ma, L., Hasan, K. M., Kramer, L. A., Zuniga, E. A., Narayana, P. A.,
et al. (2010). Diffusion tensor imaging and decision making in cocaine dependence.
PloS one, 5, e11591.

P á g i n a | 1 5 6
Lawrence, A. J., Luty, J., Bogdan, N. A., Sahakian, B. J., & Clark, L. (2009). Impulsivity and
response inhibition in alcohol dependence and problem gambling.
Psychopharmacology, 207, 163-172.
Lee, B., London, E. D., Poldrack, R. A., Farahi, J., Nacca, A., Monterosso, J. R., Mumford, J. A.,
et al. (2009). Striatal dopamine d2/d3 receptor availability is reduced in
methamphetamine dependence and is linked to impulsivity. The Journal of
neuroscience, 29, 14734-14740.
Lee, T. M. C., Zhou, W. H., Luo, X. J., Yuen, K. S. L., Ruan, X. Z., & Weng, X. C. (2005). Neural
activity associated with cognitive regulation in heroin users: A fMRI study.
Neuroscience letters, 382, 211-216.
Lee, T. M., & Pau, C. W. (2002). Impulse control differences between abstinent heroin users
and matched controls. Brain Injury, 16, 885-889.
Leland, D. S., Arce, E., Feinstein, J. S., & Paulus, M. P. (2006). Young adult stimulant users’
increased striatal activation during uncertainty is related to impulsivity. NeuroImage,
33, 725-731.
Levine, B., Dawson, D., Boutet, I., Schwartz, M. L., & Stuss, D. T. (2000). Assessment of
strategic self-regulation in traumatic brain injury: its relationship to injury severity
and psychosocial outcome. Neuropsychology, 14, 491-500.
Lezak, M. D. (2004). Neuropsychological Assessment (4th edition). New York: Oxford
University Press.
Li, C. S. R., Milivojevic, V., Kemp, K., Hong, K., & Sinha, R. (2006). Performance monitoring
and stop signal inhibition in abstinent patients with cocaine dependence. Drug and
alcohol dependence, 85, 205-212.
Li, C. S. R., & Sinha, R. (2008). Inhibitory control and emotional stress regulation:
neuroimaging evidence for frontal-limbic dysfunction in psycho-stimulant addiction.
Neuroscience and biobehavioral reviews, 32, 581-597.
Logan, G. D., Cowan, W. B., & Davis, K. A. (1984). On the ability to inhibit simple and choice
reaction time responses: a model and a method. Journal of experimental psychology.
Human perception and performance, 10, 276-291.

P á g i n a | 1 5 7
Lorenzetti, V., Lubman, D. I., Whittle, S., Solowij, N., & Yücel, M. (2010). Structural MRI
findings in long-term cannabis users: what do we know? Substance use & misuse, 45,
1787-1808.
Lubman, D. I., Yücel, M., Kettle, J. W. L., Scaffidi, A., Mackenzie, T., Simmons, J. G., & Allen, N.
B. (2009). Responsiveness to drug cues and natural rewards in opiate addiction:
associations with later heroin use. Archives of general psychiatry, 66, 205-212.
Luijten, M., Veltman, D. J., van den Brink, W., Hester, R., Field, M., Smits, M., & Franken, I. H.
A. (2011). Neurobiological substrate of smoking-related attentional bias.
NeuroImage, 54, 2374-2381.
Lundqvist, T. (2010). Imaging cognitive deficits in drug abuse. Current topics in behavioral
neurosciences, 3, 247-275.
Lyoo, I. K., Streeter, C. C., Ahn, K. H., Lee, H. K., Pollack, M. H., Silveri, M. M., Nassar, L., et al.
(2004). White matter hyperintensities in subjects with cocaine and opiate
dependence and healthy comparison subjects. Psychiatry research, 131, 135-145.
López-Torrecillas, F., Godoy García, J. F., Pérez García, M., Godoy Izquierdo, D., & Sánchez-
Barrera, M. B. (2000). Variables modulating stress and coping that discriminate drug
consumers from low or nondrug consumers. Addictive behaviors, 25, 161-165.
Madden, G. J., Petry, N. M., Badger, G. J., & Bickel, W. K. (1997). Impulsive and self-control
choices in opioid-dependent patients and non-drug-using control participants: drug
and monetary rewards. Experimental and clinical psychopharmacology, 5, 256-262.
Makris, N., Gasic, G. P., Kennedy, D. N., Hodge, S. M., Kaiser, J. R., Lee, M. J., Kim, B. W., et al.
(2008). Cortical thickness abnormalities in cocaine addiction--a reflection of both
drug use and a pre-existing disposition to drug abuse? Neuron, 60, 174-188.
Makris, N., Gasic, G. P., Seidman, L. J., Goldstein, J. M., Gastfriend, D. R., Elman, I., Albaugh, M.
D., et al. (2004b). Decreased absolute amygdala volume in cocaine addicts. Neuron,
44, 729-740.
Makris, N., Oscar-Berman, M., Jaffin, S. K., Hodge, S. M., Kennedy, D. N., Caviness, V. S.,
Marinkovic, K., et al. (2008a). Decreased volume of the brain reward system in
alcoholism. Biological psychiatry, 64, 192-202.

P á g i n a | 1 5 8
Manning, V., Wanigaratne, S., Best, D., Hill, R. G., Reed, L. J., Ball, D., Marshall, J., et al. (2008).
Changes in neuropsychological functioning during alcohol detoxification. European
addiction research, 14, 226-233.
Marinkovic, K., Oscar-Berman, M., Urban, T., O’Reilly, C. E., Howard, J. A., Sawyer, K., &
Harris, G. J. (2009). Alcoholism and dampened temporal limbic activation to
emotional faces. Alcoholism, clinical and experimental research, 33, 1880-1892.
Martinez, D., & Narendran, R. (2010). Imaging neurotransmitter release by drugs of abuse.
Current topics in behavioral neurosciences, 3, 219-245.
Martín-Santos, R., Fagundo, A. B., Crippa, J. A., Atakan, Z., Bhattacharyya, S., Allen, P., Fusar-
Poli, P., et al. (2010). Neuroimaging in cannabis use: a systematic review of the
literature. Psychological medicine, 40, 383-398.
Matochik, J. A., Eldreth, D. A., Cadet, J. L., & Bolla, K. I. (2005). Altered brain tissue
composition in heavy marijuana users. Drug and alcohol dependence, 77, 23-30.
Matochik, J. A., London, E. D., Eldreth, D. A., Cadet, J. L., & Bolla, K. I. (2003). Frontal cortical
tissue composition in abstinent cocaine abusers: a magnetic resonance imaging
study. NeuroImage, 19, 1095-1102.
Matsuo, K., Nicoletti, M., Nemoto, K., Hatch, J. P., Peluso, M. A. M., Nery, F. G., & Soares, J. C.
(2009). A voxel-based morphometry study of frontal gray matter correlates of
impulsivity. Human brain mapping, 30, 1188-1195.
Medina, K. L., Hanson, K. L., Schweinsburg, A. D., Cohen-Zion, M., Nagel, B. J., & Tapert, S. F.
(2007). Neuropsychological functioning in adolescent marijuana users: subtle deficits
detectable after a month of abstinence. Journal of the International
Neuropsychological Society, 13, 807-820.
Mintzer, M. Z., Copersino, M. L., & Stitzer, M. L. (2005). Opioid abuse and cognitive
performance. Drug and alcohol dependence, 78, 225-230.
Mishina, M. (2008). Positron emission tomography for brain research. Journal of Nihon
Medical School, 75, 68-76.
Mitchell, J. M., Fields, H. L., D’Esposito, M., & Boettiger, C. A. (2005). Impulsive responding
in alcoholics. Alcoholism, clinical and experimental research, 29, 2158-2169.

P á g i n a | 1 5 9
Miyake, A., Friedman, N. P., Emerson, M. J., Witzki, A. H., Howerter, A., & Wager, T. D.
(2000). The unity and diversity of executive functions and their contributions to
complex “Frontal Lobe” tasks: a latent variable analysis. Cognitive psychology, 41, 49-
100.
Moeller, F. G., Dougherty, D. M., Barratt, E. S., Oderinde, V., Mathias, C. W., Harper, R. A., &
Swann, A. C. (2002). Increased impulsivity in cocaine dependent subjects
independent of antisocial personality disorder and aggression. Drug and alcohol
dependence, 68, 105-111.
Moeller, F. G., Dougherty, D. M., Barratt, E. S., Schmitz, J. M., Swann, A. C., & Grabowski, J.
(2001). The impact of impulsivity on cocaine use and retention in treatment. Journal
of substance abuse treatment, 21, 193-198.
Monterosso, J. R., Aron, A. R., Cordova, X., Xu, J., & London, E. D. (2005). Deficits in response
inhibition associated with chronic methamphetamine abuse. Drug and alcohol
dependence, 79, 273-277.
Montgomery, C., Seddon, A. L., Fisk, J. E., Murphy, P. N., & Jansari, A. (2012). Cannabis-
related deficits in real-world memory. Human psychopharmacology, 27, 217-225.
Morgan, M. J., Impallomeni, L. C., Pirona, A., & Rogers, R. D. (2006). Elevated impulsivity
and impaired decision-making in abstinent Ecstasy (MDMA) users compared to
polydrug and drug-naïve controls. Neuropsychopharmacology, 31, 1562-1573.
Moriyama, Y., Muramatsu, T., Kato, M., Mimura, M., & Kashima, H. (2006). Family history of
alcoholism and cognitive recovery in subacute withdrawal. Psychiatry and clinical
neurosciences, 60, 85-89.
Nelson, S. M., Dosenbach, N. U. F., Cohen, A. L., Wheeler, M. E., Schlaggar, B. L., & Petersen, S.
E. (2010). Role of the anterior insula in task-level control and focal attention. Brain
structure & function, 214, 669-680.
Nestler, E. J. (2005). The neurobiology of cocaine addiction. Science & practice perspectives
/ a publication of the National Institute on Drug Abuse, National Institutes of Health, 3,
4-10.

P á g i n a | 1 6 0
Nestor, L. J., Ghahremani, D. G., Monterosso, J., & London, E. D. (2011). Prefrontal
hypoactivation during cognitive control in early abstinent methamphetamine-
dependent subjects. Psychiatry research, 194, 287-295.
Noël, X., Van der Linden, M., Schmidt, N., Sferrazza, R., Hanak, C., Le Bon, O., De Mol, J., et al.
(2001). Supervisory attentional system in nonamnesic alcoholic men. Archives of
general psychiatry, 58, 1152-1158.
Obrocki, J., Buchert, R., Väterlein, O., Thomasius, R., Beyer, W., & Schiemann, T. (1999).
Ecstasy--long-term effects on the human central nervous system revealed by positron
emission tomography. The British journal of psychiatry, 175, 186-188.
Obrocki, J., Schmoldt, A., Buchert, R., Andresen, B., Petersen, K., & Thomasius, R. (2002).
Specific neurotoxicity of chronic use of ecstasy. Toxicology letters, 127, 285-297.
Observatorio Español sobre Drogas (2009). Informe 2009. Descargado de:
http://www.pnsd.msc.es/Categoria2/observa/pdf/oed-2009.pdf el 2 de Mayo de
2012.
Oquendo, M. A., Baca-García, E., Graver, R., Morales, M., Montalvan, V., & Mann, J. J. (2001).
Spanish adaptation of the Barratt Impulsiveness Scale (BIS-11). European Journal of
Psychiatry, 15, 147-155.
Pace-Schott, E. F., Morgan, P. T., Malison, R. T., Hart, C. L., Edgar, C., Walker, M., & Stickgold,
R. (2008). Cocaine users differ from normals on cognitive tasks which show poorer
performance during drug abstinence. The American journal of drug and alcohol abuse,
34, 109-121.
Parada, M., Corral, M., Mota, N., Crego, A., Rodríguez Holguín, S., & Cadaveira, F. (2012).
Executive functioning and alcohol binge drinking in university students. Addictive
behaviors, 37, 167-172.
Parrott, A. C., Sisk, E., & Turner, J. J. (2000). Psychobiological problems in heavy “ecstasy”
(MDMA) polydrug users. Drug and alcohol dependence, 60, 105-110.
Parrott, A. C. (2012). MDMA and serotonergic neurotoxicity: the empirical evidence for its
adverse effects in humans-no need for translation. British journal of pharmacology, en
prensa.

P á g i n a | 1 6 1
Passetti, F., Clark, L., Mehta, M. A., Joyce, E., & King, M. (2008). Neuropsychological
predictors of clinical outcome in opiate addiction. Drug and alcohol dependence, 94,
82-91.
Patton, J. H., Stanford, M. S., & Barratt, E. S. (1995). Factor structure of the Barratt
impulsiveness scale. Journal of clinical psychology, 51, 768-774.
Paulus, M. P., Tapert, S. F., & Schuckit, M. A. (2005). Neural activation patterns of
methamphetamine-dependent subjects during decision making predict relapse.
Archives of general psychiatry, 62, 761-768.
Pérez, J., & Torrubia, R. (1986). Fiabilidad y validez de la versión española de la escala de
búsqueda de sensaciones (formaV). Revista Latinoamericana de Psicología, 18, 7-22.
Pertwee, R. G., & Ross, R. A. (2002). Cannabinoid receptors and their ligands.
Prostaglandins, leukotrienes, and essential fatty acids, 66, 101-121.
Petry, N. M. (2001). Delay discounting of money and alcohol in actively using alcoholics,
currently abstinent alcoholics, and controls. Psychopharmacology, 154, 243-250.
Pezawas, L., Fischer, G., Podreka, I., Schindler, S., Brücke, T., Jagsch, R., Thurnher, M., et al.
(2002). Opioid addiction changes cerebral blood flow symmetry. Neuropsychobiology,
45, 67-73.
Pezawas, L. M., Fischer, G., Diamant, K., Schneider, C., Schindler, S. D., Thurnher, M., Ploechl,
W., et al. (1998). Cerebral CT findings in male opioid-dependent patients:
stereological, planimetric and linear measurements. Psychiatry research, 83, 139-147.
Pillay, S. S., Rogowska, J., Kanayama, G., Gruber, S., Simpson, N., Pope, H. G., & Yurgelun-
Todd, D. A. (2008). Cannabis and motor function: fMRI changes following 28 days of
discontinuation. Experimental and clinical psychopharmacology, 16, 22-32.
Pitel, A. L., Perruchet, P., Vabret, F., Desgranges, B., Eustache, F., & Beaunieux, H. (2010).
The advantage of errorless learning for the acquisition of new concepts’ labels in
alcoholics. Psychological medicine, 40, 497-502.
Pope, H. G., Gruber, A. J., Hudson, J. I., Cohane, G., Huestis, M. A., & Yurgelun-Todd, D.
(2003). Early-onset cannabis use and cognitive deficits: what is the nature of the
association? Drug and alcohol dependence, 69, 303-310.

P á g i n a | 1 6 2
Posner, M. I., Rothbart, M. K., Sheese, B. E., & Tang, Y. (2007). The anterior cingulate gyrus
and the mechanism of self-regulation. Cognitive, affective & behavioral neuroscience,
7, 391-395.
Potenza, M. N. (2001). The neurobiology of pathological gambling. Seminars in clinical
neuropsychiatry, 6, 217-226.
Prince van Leeuwen, A., Creemers, H. E., Verhulst, F. C., Ormel, J., & Huizink, A. C. (2011).
Are adolescents gambling with cannabis use? A longitudinal study of impulsivity
measures and adolescent substance use: the TRAILS study. Journal of studies on
alcohol and drugs, 72, 70-78.
Prosser, J. M., Eisenberg, D., Davey, E. E., Steinfeld, M., Cohen, L. J., London, E. D., &
Galynker, I. I. (2008). Character pathology and neuropsychological test performance
in remitted opiate dependence. Substance abuse treatment, prevention, and policy, 3,
23.
Quednow, B. B., Kühn, K. U., Hoppe, C., Westheide, J., Maier, W., Daum, I., & Wagner, M.
(2007). Elevated impulsivity and impaired decision-making cognition in heavy users
of MDMA (“Ecstasy”). Psychopharmacology, 189, 517-530.
Redish, A. D., Jensen, S., & Johnson, A. (2008). A unified framework for addiction:
vulnerabilities in the decision process. The Behavioral and brain sciences, 31, 415-
437.
Reid, A. G., Daglish, M. R. C., Kempton, M. J., Williams, T. M., Watson, B., Nutt, D. J., &
Lingford-Hughes, A. R. (2008). Reduced thalamic grey matter volume in opioid
dependence is influenced by degree of alcohol use: a voxel-based morphometry
study. Journal of psychopharmacology, 22, 7-10.
Rey, A. (1941). L'examen psychologique dans les cas d'encephalopathie traumatique.
Archives de Psychologie, 28, 286-340.
Reynolds, B. (2006). A review of delay-discounting research with humans: relations to
drug use and gambling. Behavioural pharmacology, 17, 651-667.
Reynolds, B., & Schiffbauer, R. (2004). Measuring state changes in human delay
discounting: an experiential discounting task. Behavioural processes, 67, 343-356.

P á g i n a | 1 6 3
Rogers, R. D., Everitt, B. J., Baldacchino, A., Blackshaw, A. J., Swainson, R., Wynne, K., Baker,
N. B., et al. (1999a). Dissociable deficits in the decision-making cognition of chronic
amphetamine abusers, opiate abusers, patients with focal damage to prefrontal
cortex, and tryptophan-depleted normal volunteers: evidence for monoaminergic
mechanisms. Neuropsychopharmacology, 20, 322-339.
Rogers, R. D., Owen, A. M., Middleton, H. C., Williams, E. J., Pickard, J. D., Sahakian, B. J., &
Robbins, T. W. (1999b). Choosing between small, likely rewards and large, unlikely
rewards activates inferior and orbital prefrontal cortex. The Journal of neuroscience,
19, 9029-9038.
Rogers, R. D., & Robbins, T. W. (2001). Investigating the neurocognitive deficits associated
with chronic drug misuse. Current opinion in neurobiology, 11, 250-257.
Roberts, A. C., Robbins, T. W., & Weiskrantz, L. (1998). The Prefrontal Cortex: Executive and
Cognitive Functions. New York: Oxford University Press.
Romero, M. J., Asensio, S., Palau, C., Sanchez, A., & Romero, F. J. (2010). Cocaine addiction:
diffusion tensor imaging study of the inferior frontal and anterior cingulate white
matter. Psychiatry research, 181, 57-63.
Rose, J. E., Behm, F. M., Westman, E. C., Mathew, R. J., London, E. D., Hawk, T. C., Turkington,
T. G., et al. (2003). PET studies of the influences of nicotine on neural systems in
cigarette smokers. The American journal of psychiatry, 160, 323-333.
Ruff, R. M. (1996). Ruff Figural Fluency Test: Professional Manual. Lutz, Florida:
Psychological Assessment Resources.
Salloum, J. B., Ramchandani, V. A., Bodurka, J., Rawlings, R., Momenan, R., George, D., &
Hommer, D. W. (2007). Blunted rostral anterior cingulate response during a
simplified decoding task of negative emotional facial expressions in alcoholic
patients. Alcoholism, clinical and experimental research, 31, 1490-1504.
Salthouse TA (2009) When does age-related cognitive decline begin? Neurobiology of
Aging, 30, 507-514.
Schulz, S. (2011). MDMA & cannabis: a mini-review of cognitive, behavioral, and
neurobiological effects of co-consumption. Current drug abuse reviews, 4, 81-86.

P á g i n a | 1 6 4
Sedó, M. (2005). Test de los Cinco Dígitos: Five Digit Test. Madrid: TEA Ediciones.
Sevy, S., Smith, G. S., Ma, Y., Dhawan, V., Chaly, T., Kingsley, P. B., Kumra, S., et al. (2008).
Cerebral glucose metabolism and D2/D3 receptor availability in young adults with
cannabis dependence measured with positron emission tomography.
Psychopharmacology, 197, 549-556.
Sim, M. E., Lyoo, I. K., Streeter, C. C., Covell, J., Sarid-Segal, O., Ciraulo, D. A., Kim, M. J., et al.
(2007). Cerebellar gray matter volume correlates with duration of cocaine use in
cocaine-dependent subjects. Neuropsychopharmacology, 32, 2229-2237.
Sinha, R., & Li, C. S. R. (2007). Imaging stress- and cue-induced drug and alcohol craving:
association with relapse and clinical implications. Drug and alcohol review, 26, 25-31.
Smith, G. T., Fischer, S., Cyders, M. A., Annus, A. M., Spillane, N. S., & McCarthy, D. M. (2007).
On the validity and utility of discriminating among impulsivity-like traits. Assessment,
14, 155-170.
Sofuoglu, M. (2010). Cognitive enhancement as a pharmacotherapy target for stimulant
addiction. Addiction, 105, 38-48.
Spiro, R. J., & Jehng, J. (1990). Cognitive flexibility and hipertext: Theory and technology for
the non-linear and multidimensional traversal of complex subject matter. D. Nix y R.
Spiro (Eds.), Cognition, Education, and Multimedia. Hillsdale, New Jersey: Erlbaum.
Squeglia, L. M., Sorg, S. F., Schweinsburg, A. D., Wetherill, R. R., Pulido, C., & Tapert, S. F.
(2012). Binge drinking differentially affects adolescent male and female brain
morphometry. Psychopharmacology, 220, 529-539.
Stout, J. C., Busemeyer, J. R., Lin, A., Grant, S. J., & Bonson, K. R. (2004). Cognitive modeling
analysis of decision-making processes in cocaine abusers. Psychonomic bulletin &
review, 11, 742-747.
Streeter, C. C., Terhune, D. B., Whitfield, T. H., Gruber, S., Sarid-Segal, O., Silveri, M. M.,
Tzilos, G., et al. (2008). Performance on the Stroop predicts treatment compliance in
cocaine-dependent individuals. Neuropsychopharmacology, 33, 827-836.
Stuss, D. T., & Alexander, M. P. (2000). Executive functions and the frontal lobes: a
conceptual view. Psychological Research, 63, 289-298.

P á g i n a | 1 6 5
Stuss, D. T., & Knight, R. T. (2002). Principles of frontal lobe functioning. New York: Oxford
University Press.
Sullivan, E. V., Deshmukh, A., De Rosa, E., Rosenbloom, M. J., & Pfefferbaum, A. (2005).
Striatal and forebrain nuclei volumes: contribution to motor function and working
memory deficits in alcoholism. Biological psychiatry, 57, 768-776.
Tanabe, J., Tregellas, J. R., Dalwani, M., Thompson, L., Owens, E., Crowley, T., & Banich, M.
(2009). Medial orbitofrontal cortex gray matter is reduced in abstinent substance-
dependent individuals. Biological psychiatry, 65, 160-164.
Tapert, S. F., Brown, G. G., Baratta, M. V., & Brown, S. A. (2004). fMRI BOLD response to
alcohol stimuli in alcohol dependent young women. Addictive behaviors, 29, 33-50.
Tapert, S. F., Schweinsburg, A. D., Drummond, S. P. A., Paulus, M. P., Brown, S. A., Yang, T. T.,
& Frank, L. R. (2007). Functional MRI of inhibitory processing in abstinent adolescent
marijuana users. Psychopharmacology, 194, 173-183.
Tedstone, D., & Coyle, K. (2004). Cognitive impairments in sober alcoholics: performance
on selective and divided attention tasks. Drug and alcohol dependence, 75, 277-286.
Thomas, L. A., Hall, J. M., Skup, M., Jenkins, S. E., Pine, D. S., & Leibenluft, E. (2011). A
developmental neuroimaging investigation of the change paradigm. Developmental
science, 14, 148-161.
Tomasi, D., Goldstein, R. Z., Telang, F., Maloney, T., Alia-Klein, N., Caparelli, E. C., & Volkow,
N. D. (2007). Widespread disruption in brain activation patterns to a working
memory task during cocaine abstinence. Brain research, 1171, 83-92.
Torrubia, R., Ávila, C., Moltó, J., & Caseras, X. (2001). The Sensitivity to Punishment and
Sensitivity to Reward Questionnaire (SPSRQ) as a measure of Gray’s anxiety and
impulsivity dimensions. Personality and Individual Differences, 31, 837-862.
Townshend, J. M., & Duka, T. (2003). Mixed emotions: alcoholics’ impairments in the
recognition of specific emotional facial expressions. Neuropsychologia, 41, 773-782.
Tucker, K. A., Potenza, M. N., Beauvais, J. E., Browndyke, J. N., Gottschalk, P. C., & Kosten, T.
R. (2004). Perfusion abnormalities and decision making in cocaine dependence.
Biological psychiatry, 56, 527-530.

P á g i n a | 1 6 6
Turner, T. H., LaRowe, S., Horner, M. D., Herron, J., & Malcolm, R. (2009). Measures of
cognitive functioning as predictors of treatment outcome for cocaine dependence.
Journal of substance abuse treatment, 37, 328-334.
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello, F., Etard, O., Delcroix, N.,
Mazoyer, B., et al. (2002). Automated anatomical labeling of activations in SPM using
a macroscopic anatomical parcellation of the MNI MRI single-subject brain.
NeuroImage, 15, 273-289.
United Nations Office on Drugs and Crime (2011). United Nations World Drug Report.
Descargado de: http://www.unodc.org/unodc/en/data-and-analysis/WDR-
2011.html el 2 de Mayo de 2012.
van Holst, R. J., & Schilt, T. (2011). Drug-related decrease in neuropsychological functions
of abstinent drug users. Current drug abuse reviews, 4, 42-56.
van Holst, R. J., de Ruiter, M. B., van den Brink, W., Veltman, D. J., & Goudriaan, A. E. (2012).
A voxel-based morphometry study comparing problem gamblers, alcohol abusers,
and healthy controls. Drug and alcohol dependence, en prensa.
Verdejo, A., Toribio, I., Orozco, C., Puente, K. L., & Pérez-García, M. (2005).
Neuropsychological functioning in methadone maintenance patients versus abstinent
heroin abusers. Drug and alcohol dependence, 78, 283-288.
Verdejo-García, A., Benbrook, A., Funderburk, F., David, P., Cadet, J.-L., & Bolla, K. I. (2007a).
The differential relationship between cocaine use and marijuana use on decision-
making performance over repeat testing with the Iowa Gambling Task. Drug and
alcohol dependence, 90, 2-11.
Verdejo-García, A, Betanzos-Espinosa, P., Lozano, O. M., Vergara-Moragues, E., González-
Saiz, F., Fernández-Calderón, F., Bilbao-Acedos, I., et al. (2012). Self-regulation and
treatment retention in cocaine dependent individuals: a longitudinal study. Drug and
alcohol dependence, 122, 142-148.
Verdejo-García, A., & Bechara, A. (2009). A somatic marker theory of addiction.
Neuropharmacology, 56, 48-62.
Verdejo-García, A., Bechara, A., Recknor, E. C., & Pérez-García, M. (2006). Executive
dysfunction in substance dependent individuals during drug use and abstinence: an

P á g i n a | 1 6 7
examination of the behavioral, cognitive and emotional correlates of addiction.
Journal of the International Neuropsychological Society, 12, 405-415.
Verdejo-García, A., Bechara, A., Recknor, E. C., & Pérez-García, M. (2007c). Negative
emotion-driven impulsivity predicts substance dependence problems. Drug and
alcohol dependence, 91, 213-219.
Verdejo-García, A. J., López-Torrecillas, F., Aguilar de Arcos, F., & Pérez-García, M. (2005).
Differential effects of MDMA, cocaine, and cannabis use severity on distinctive
components of the executive functions in polysubstance users: a multiple regression
analysis. Addictive behaviors, 30, 89-101.
Verdejo-García, A. J., Perales, J. C., & Pérez-García, M. (2007b). Cognitive impulsivity in
cocaine and heroin polysubstance abusers. Addictive behaviors, 32, 950-966.
Verdejo-García, A., Lawrence, A. J., & Clark, L. (2008). Impulsivity as a vulnerability marker
for substance-use disorders: review of findings from high-risk research, problem
gamblers and genetic association studies. Neuroscience and biobehavioral reviews, 32,
777-810.
Verdejo-García, A., Lozano, O., Moya, M., Alcázar, M. A., & Pérez-García, M. (2010).
Psychometric properties of a Spanish version of the UPPS-P impulsive behavior scale:
reliability, validity and association with trait and cognitive impulsivity. Journal of
personality assessment, 92, 70-77.
Verdejo-García, A., López-Torrecillas, F., Giménez, C. O., & Pérez-García, M. (2004). Clinical
implications and methodological challenges in the study of the neuropsychological
correlates of cannabis, stimulant, and opioid abuse. Neuropsychology review, 14, 1-41.
Verdejo-García, A., & Pérez-García, M. (2007). Profile of executive deficits in cocaine and
heroin polysubstance users: common and differential effects on separate executive
components. Psychopharmacology, 190, 517-530.
Verdejo-García, A., Rivas-Pérez, C., Vilar-López, R., & Pérez-García, M. (2007d). Strategic
self-regulation, decision-making and emotion processing in poly-substance abusers
in their first year of abstinence. Drug and alcohol dependence, 86, 139-146.
Volkow, N. D., Chang, L., Wang, G. J., Fowler, J. S., Ding, Y. S., Sedler, M., Logan, J., et al.
(2001). Low level of brain dopamine D2 receptors in methamphetamine abusers:

P á g i n a | 1 6 8
association with metabolism in the orbitofrontal cortex. The American journal of
psychiatry, 158, 2015-2021.
Volkow, N. D., Fowler, J. S., Wang, G. J., Hitzemann, R., Logan, J., Schlyer, D. J., Dewey, S. L., et
al. (1993). Decreased dopamine D2 receptor availability is associated with reduced
frontal metabolism in cocaine abusers. Synapse, 14, 169-177.
Volkow, N. D., Gillespie, H., Mullani, N., Tancredi, L., Grant, C., Valentine, A., & Hollister, L.
(1996). Brain glucose metabolism in chronic marijuana users at baseline and during
marijuana intoxication. Psychiatry research, 67, 29-38.
Volkow, N. D., Hitzemann, R., Wang, G. J., Fowler, J. S., Wolf, A. P., Dewey, S. L., &
Handlesman, L. (1992). Long-term frontal brain metabolic changes in cocaine
abusers. Synapse, 11, 184-190.
Volkow, N. D., Ma, Y., Zhu, W., Fowler, J. S., Li, J., Rao, M., Mueller, K., et al. (2008). Moderate
doses of alcohol disrupt the functional organization of the human brain. Psychiatry
research, 162, 205-213.
Volkow, N. D., Tomasi, D., Wang, G. J., Fowler, J. S., Telang, F., Goldstein, R. Z., Alia-Klein, N.,
et al. (2011). Reduced metabolism in brain “control networks” following cocaine-cues
exposure in female cocaine abusers. PloS one, 6, e16573.
Volkow, N. D., Fowler, J. S., Wang, G. J., Telang, F., Logan, J., Jayne, M., Ma, Y., et al. (2010).
Cognitive control of drug craving inhibits brain reward regions in cocaine abusers.
NeuroImage, 49, 2536-2543.
Volkow, N. D., Wang, G. J., Telang, F., Fowler, J. S., Logan, J., Childress, A. R., Jayne, M., et al.
(2006). Cocaine cues and dopamine in dorsal striatum: mechanism of craving in
cocaine addiction. The Journal of neuroscience, 26, 6583-6588.
Volz, K. G., Schubotz, R. I., & von Cramon, D. Y. (2003). Predicting events of varying
probability: uncertainty investigated by fMRI. NeuroImage, 19, 271-280.
Wang, G. J., Volkow, N. D., Fowler, J. S., Cervany, P., Hitzemann, R. J., Pappas, N. R., Wong, C.
T., et al. (1999). Regional brain metabolic activation during craving elicited by recall
of previous drug experiences. Life sciences, 64, 775-784.

P á g i n a | 1 6 9
Wang, Z. X., Zhang, J. X., Wu, Q.-L., Liu, N., Hu, X.-P., Chan, R. C. K., & Xiao, Z. W. (2010).
Alterations in the processing of non-drug-related affective stimuli in abstinent heroin
addicts. NeuroImage, 49, 971-976.
Watter, S., Geffen, G. M., & Geffen, L. B. (2001). The n-back as a dual-task: P300 morphology
under divided attention. Psychophysiology, 38, 998-1003.
Weijers, H. G., Wiesbeck, G. A., & Böning, J. (2001). Reflection–impulsivity, personality and
performance: a psychometric and validity study of the Matching Familiar Figures
Test in detoxified alcoholics. Personality and Individual Differences, 31, 731-754.
Wechsler, D. (1997). Wechsler memory scale (3rd Edition). San Antonio, Texas: The
Psychological Corporation.
Whiteside, S. P., & Lynam, D. R. (2001). The five factor model and impulsivity: using a
structural model of personality to understand impulsivity. Personality and Individual
Differences, 30, 669-689.
Whiteside, S. P., & Lynam, D. R. (2003). Understanding the role of impulsivity and
externalizing psychopathology in alcohol abuse: application of the UPPS impulsive
behavior scale. Experimental and Clinical Psychopharmacology, 11, 210-217.
Wilson, W., Mathew, R., Turkington, T., Hawk, T., Coleman, R. E., & Provenzale, J. (2000).
Brain morphological changes and early marijuana use: a magnetic resonance and
positron emission tomography study. Journal of addictive diseases, 19, 1-22.
Woicik, P. A., Moeller, S. J., Alia-Klein, N., Maloney, T., Lukasik, T. M., Yeliosof, O., Wang, G.-J.,
et al. (2009). The neuropsychology of cocaine addiction: recent cocaine use masks
impairment. Neuropsychopharmacology, 34, 1112-1122.
Yaouhi, K., Bertran, F., Clochon, P., Mézenge, F., Denise, P., Foret, J., Eustache, F., et al.
(2009). A combined neuropsychological and brain imaging study of obstructive sleep
apnea. Journal of sleep research, 18, 36-48.
Yip, J. T., & Lee, T.M. (2006). Selective impairment of sadness and disgust recognition in
abstinent ecstasy users. Neuropsychologia, 44, 959-965.

P á g i n a | 1 7 0
Young, A. W., Perrett, D. I., Calder, A. J., Sprengelmeyer, R., & Ekman, P. (2002). Facial
expressions of emotion: stimuli and tests (FEEST). Bury St. Edmunds: Thames Valley
Test Company.
Yuan, Y., Zhu, Z., Shi, J., Zou, Z., Yuan, F., Liu, Y., Lee, T. M. C., et al. (2009). Gray matter
density negatively correlates with duration of heroin use in young lifetime heroin-
dependent individuals. Brain and cognition, 71, 223-228.
Yücel, M., Solowij, N., Respondek, C., Whittle, S., Fornito, A., Pantelis, C., & Lubman, D. I.
(2008). Regional brain abnormalities associated with long-term heavy cannabis use.
Archives of general psychiatry, 65, 694-701.
Zakzanis, K. K. (2001). Statistics to tell the truth, the whole truth, and nothing but the
truth: formulae, illustrative numerical examples, and heuristic interpretation of effect
size analyses for neuropsychological researchers. Archives of clinical neuropsychology,
16, 653-667.
Zakzanis, K. K., Campbell, Z., & Jovanovski, D. (2007). The neuropsychology of ecstasy
(MDMA) use: a quantitative review. Human psychopharmacology, 22, 427-435.
Zinn, S., Stein, R., & Swartzwelder, H. S. (2004). Executive functioning early in abstinence
from alcohol. Alcoholism, clinical and experimental research, 28, 1338-1346.
Zubieta, J. K., Heitzeg, M. M., Xu, Y., Koeppe, R. A., Ni, L., Guthrie, S., & Domino, E. F. (2005).
Regional cerebral blood flow responses to smoking in tobacco smokers after
overnight abstinence. The American journal of psychiatry, 162, 567-577.
Zuckerman, M., Kuhlman, D., Joireman, J., Teta, P., & Kraft, M. (1993). A comparison of the
three structural models for the personality: the big three, the big five and the
alternative five. Journal of Personality and Social Psychology, 65, 747-768.

P á g i n a | 1 7 1
[ANEXOS]

[ANEXO I]

G
D
Ti
LEa
b
c
d
e
a
ARRAA
KCIGWVU
1
lfHcuitec
p
0d
ARTICLE IN PRESS Model
AD-4380; No. of Pages 7
Drug and Alcohol Dependence xxx (2012) xxx– xxx
Contents lists available at SciVerse ScienceDirect
Drug and Alcohol Dependence
jo u rn al hom epage: www.elsev ier .com/ locate /drugalcdep
rait impulsivity and prefrontal gray matter reductions in cocaine dependentndividuals
aura Moreno-Lópeza, Andrés Catenab, María José Fernández-Serranoc, Elena Delgado-Ricoa,mmanuel A. Stamatakisd, Miguel Pérez-Garcíaa,e, Antonio Verdejo-Garcíaa,e,∗
Department of Clinical Psychology, School of Psychology, University of Granada, SpainDepartment of Experimental Psychology and Behavioral Physiology, School of Psychology, University of Granada, SpainDepartment of Psychology, University of Jaen, SpainDivision of Anaesthesia, School of Clinical Medicine, University of Cambridge, UKInstitute of Neuroscience F. Olóriz, University of Granada, Spain
r t i c l e i n f o
rticle history:eceived 22 August 2011eceived in revised form 9 February 2012ccepted 10 February 2012vailable online xxx
eywords:ocaine
mpulsivityray matterhite matter
oxel based morphometryPPS
a b s t r a c t
Background: Impulsivity is thought to play a key role in cocaine addiction onset and progression; therefore,we hypothesized that different facets of impulsive personality may be significantly associated with brainstructural abnormalities in cocaine-dependent individuals.Methods: Thirty-eight cocaine-dependent individuals and 38 non-drug using controls completed theUPPS-P scale (measuring five different facets of impulsivity: sensation seeking, lack of premeditation,lack of perseverance, and positive and negative urgency) and were scanned on a 3T MRI scanner. Weused whole-brain voxel-based morphometry analyses (VBM) to detect differences in gray matter (GM)and white matter (WM) volumes between cocaine users and controls, and to measure differences in theway that impulsivity relates to GM and WM volumes in cocaine users vs. controls.Results: Cocaine-dependent individuals had lower GM volumes in a number of sections of the orbitofrontalcortex, right inferior frontal gyrus, right insula, left amygdala and parahippocampal gyrus, temporalgyrus, and bilateral caudate. They also had lower WM volumes in the left inferior and medial frontal
gyrus, superior temporal gyrus, right anterior cingulate cortex, insula and caudate. There was a positivecorrelation between trait impulsivity and GM volume in the left inferior/middle frontal gyrus of cocaine-dependent individuals, a pattern directly opposed to the association in controls. Conversely, in cocaineusers lack of premeditation was negatively correlated with GM volume in the insula and the putamen.Conclusions: Trait impulsivity may influence cocaine dependence by impacting its neurobiological under-pinnings in frontostriatal systems.. Introduction
Cocaine addiction is a worldwide major public health prob-em for which current prevention and treatment options are notully satisfactory (EMCDDA, 2009; Substance Abuse and Mentalealth Services Administration, 2010). Neuroimaging studies inocaine-dependent individuals have revealed significant brain vol-me reductions, most notably in a priori selected regions of
nterest, such as the striatum, the amygdala or the prefrontal cor-
Please cite this article in press as: Moreno-López, L., et al., Trait impulsivity anDrug Alcohol Depend. (2012), doi:10.1016/j.drugalcdep.2012.02.012
ex (Barrós-Loscertales et al., 2011; Makris et al., 2004; Matochikt al., 2003; Tanabe et al., 2009). These regions are thought toontribute to critical aspects of the addictive cycle, including
∗ Corresponding author at: Facultad de Psicología, Universidad de Granada Cam-us de Cartuja S/N, 18071 Granada, Spain. Tel.: +34 958242948; fax: +34 958243749.
E-mail address: [email protected] (A. Verdejo-García).
376-8716/$ – see front matter © 2012 Elsevier Ireland Ltd. All rights reserved.oi:10.1016/j.drugalcdep.2012.02.012
© 2012 Elsevier Ireland Ltd. All rights reserved.
reinforcement learning, craving, and inhibitory control (Koob andVolkow, 2010). Nonetheless, the findings from nonbiased auto-mated techniques, such as Voxel Based Morphometry (VBM),have yielded inconsistent results. Despite previous positivefindings showing significant gray matter (GM) reductions incocaine-dependent individuals, including the medial prefrontalcortex, superior temporal cortex, insula, thalamus and cerebel-lum (Franklin et al., 2002; Sim et al., 2007), a recent VBM studyfailed to detect any structural change (GM or white matter (WM)reductions) in cocaine users compared to non-drug using controls(Narayana et al., 2010). Perhaps more critically, most of these stud-ies have failed to find any correlation between estimates of druguse patterns (e.g. amount, duration or age at onset) and GM and
d prefrontal gray matter reductions in cocaine dependent individuals.
WM reductions (Franklin et al., 2002; Makris et al., 2004; Matochiket al., 2003; but see Ersche et al., 2011). This apparent lack ofassociation between cocaine exposure and brain attrition raisesthe possibility that certain personality traits, such as impulsivity,

ING Model
D
2 Alcoho
mi
mtetrthStisvittastftcsii
yccduiuldragftaeas
2
2
npdprt(httatccdd(
ARTICLEAD-4380; No. of Pages 7
L. Moreno-López et al. / Drug and
ay relate to GM and WM abnormalities in cocaine-dependentndividuals.
Impulsivity is viewed as a multifaceted trait that varies nor-ally across the population, but that in high levels may predispose
o a range of dysfunctional behaviors, including addiction (Cyderst al., 2007; Verdejo-García et al., 2008). Animal studies have shownhat increased impulsivity is associated with reduced dopamineeceptors availability and cocaine use escalation and progressiono dependence (Belin et al., 2008; Dalley et al., 2007). In humans,igh levels of trait impulsivity (indexed with the Barratt Impulsivitycale) are associated with lower midbrain dopamine autorecep-or binding, and greater amphetamine-induced dopamine releasen the striatum (Buckholtz et al., 2010). In addition, trait impul-ivity is negatively correlated with orbitofrontal cortex (OFC) GMolumes in healthy individuals (Matsuo et al., 2009). Nonetheless,n addicted individuals, continued use of stimulants is thoughto further exacerbate impulsive traits (Ersche et al., 2010), ando possibly modify its neural underpinnings. In fact, the associ-tion between trait impulsivity levels and striatal dysfunction istronger in methamphetamine-dependent individuals comparedo healthy controls (Lee et al., 2009). Conversely, Ersche et al. (2011)ailed to find significant correlations between particular aspects ofrait impulsivity (impulsive reward-seeking) and GM reductions inocaine-dependent subjects. Therefore, more studies are needed topecifically explore if, as expected, different facets of trait impulsiv-ty are specifically linked to GM volumes in frontostriatal regionsn cocaine-dependent individuals.
The aims of this study were (i) to use whole-brain VBM anal-ses to examine possible GM and WM reductions in a sample ofurrently abstinent (>1 month) cocaine-dependent individuals, asompared to a non-drug using control group; and (ii) to measureifferences in the way impulsivity relates to GM and WM vol-mes in cocaine users vs. controls. Because impulsive personality
n the normal population is negatively associated with GM vol-me in the OFC (Matsuo et al., 2009), which is also impacted by
ifetime cocaine use (Alia-Klein et al., 2011), we expected cocaine-ependent individuals to have reduced GM volumes in the OFC andichly interconnected regions, such as insula, amygdala, striatum,nd WM adjacent to these regions (Bechara, 2005). Because pro-ression of cocaine use is thought to provoke neuroadaptationsrom the ventral striatum to the dorsal striatum, and from the OFCo more extensive prefrontal regions like the anterior cingulatend dorsolateral prefrontal cortex (Koob and Volkow, 2010), wexpected impulsivity to be uniquely associated with more lateralspects of the prefrontal cortex and more posterior aspects of thetriatum in cocaine-dependent individuals.
. Materials and methods
.1. Participants
Thirty-eight cocaine-dependent individuals, mean age = 29.58, SD = 6.53, and 38on-drug using controls, mean age = 31.08, SD = 5.14, participated in this study. Allarticipants were male due to the low prevalence of women entering drug treatmenturing the recruitment period. Cocaine users were recruited in an inpatient thera-eutic community (“Proyecto Hombre”), in the city of Granada, Spain. All of themeported cocaine as their main drug of choice and the one for which they requestedreatment. Clinical interviews based on Diagnostic and Statistical Manual version IVDSM-IV) criteria confirmed cocaine dependence diagnosis; nonetheless, they alsoad regular use of tobacco, alcohol, cannabis and MDMA (see Table 1). To enterhe study, cocaine users had to be abstinent for at least 30 days (for any drug butobacco), as confirmed by weekly urine tests; in this way, we could rule out acutend residual effects of previously used drugs on brain structure, with the excep-ion of nicotine (84.2% of cocaine-dependent individuals and 44.7% of controls were
Please cite this article in press as: Moreno-López, L., et al., Trait impulsivity anDrug Alcohol Depend. (2012), doi:10.1016/j.drugalcdep.2012.02.012
urrent smokers). None of the cocaine patients were currently following pharma-ological substitution treatments. Potential participants who had previously beeniagnosed with any disorder from DSM-IV Axes I and II (other than substance depen-ence), or had neurological or systemic diseases affecting central nervous systemCNS) functioning were excluded.
PRESSl Dependence xxx (2012) xxx– xxx
Non-drug using controls were recruited through adverts distributed by a localemployment agency, and therefore they were also matched to cocaine participantsin terms of unemployment status. Selection criteria for control participants were:(i) absence of current or past substance use, excluding past or current social drink-ing (less than ten standard alcohol units per week); (ii) absence of documentedmajor psychiatric disorders; (iii) absence of documented head injury or neurologicaldisorder, and (iv) not being taking medication with effects on the CNS.
For both groups, evidence of stroke or space-occupying lesions observed onconventional clinical MR images, any contraindications to MRI scanning (includingclaustrophobia and implanted ferromagnetic objects), and history of loss of con-sciousness (LOC) for longer than 30 min or LOC with any neurological consequencewere exclusionary.
2.2. Instruments and assessment procedures
The study was approved by the Ethics Committee for Research in Humans of theUniversity of Granada. All participants signed an informed consent form certifyingtheir voluntary participation. Controls, but not patients, received a D 40 compensa-tion for participating in the study. Assessments were conducted across two sessionsseparated by less than one week. During the first session we administered the Inter-view for Research on Addictive Behavior, the Hamilton Rating Scales for Depressionand Anxiety (HAM-D and HAM-A), and the UPPS-P Impulsive Behavior Scale, alongwith a battery of cognitive tests of which results will be reported separately. Thesecond session involved the MRI scanning. The MRI scans lasted approximately6 min.
2.2.1. Patterns of drug use. Data regarding lifetime amount and duration of use ofthe different drugs was self-reported by participants and collected using the Inter-view for Research on Addictive Behavior (Verdejo-García et al., 2005). This interviewprovides an estimation of monthly use of each substance during regular use (amountper month) and total duration of use of each substance (in years). The descriptivescores for these variables in the sample are presented in Table 1.
2.2.2. Trait impulsivity. We used the Spanish version of the UPPS-P Scale (Verdejo-García et al., 2010; Whiteside and Lynam, 2001). This is a 59-item self-reportinventory designed to measure five distinct personality pathways to impulsivebehavior: sensation seeking, (lack of) perseverance, (lack of) premeditation, neg-ative urgency, and positive urgency. The first 4 dimensions were included in theoriginal version of the UPPS scale (Whiteside and Lynam, 2001); the fifth dimensionhas been included based on recent work by (Cyders et al., 2007; Smith et al., 2007).Sensation seeking (12 items) incorporates two aspects: (1) a tendency to enjoy andpursue activities that are exciting, and (2) an openness to trying new experiencesthat may or may not be dangerous; (lack of) perseverance (10 items) refers to theindividual’s ability to remain focused on a task that may be boring or difficult; (lackof) premeditation (11 items) refers to the tendency to think and reflect on the con-sequences of an act before engaging in that act; and finally urgency (12 items) refersto the tendency to experience strong impulses under conditions of negative affect(negative urgency, 12 items) or positive affect (positive urgency, 14 items). Each itemon the UPPS is rated on a 4-point scale ranging from 1 (strongly agree) to 4 (stronglydisagree). We obtained the total scores of each of these five UPPS-P dimensions foranalysis (Table 2).
2.3. MRI acquisition
Participants were scanned on a 3T whole body MRI scanner (Phillips Achieva)operating with 8 channels phased-array head coil for reception. For each participant,a T1-weighted 3D volume was acquired using a T1-weighted 3D-turbo-gradient-echo sequence (3D-TFE), in sagittal orientation with 0.94 × 0.94 × 1.0 mm resolution(160 slides, FOV = 240 × 240 mm2, matrix 256 × 256 × 160) with repetition time of8 ms, echo time 4 ms, inversion delay = 1022.6264 ms, flip angle of 8◦ , band with191 Hz/pixel. The sequence was optimal for reducing motion sensitivity, suscepti-bility artifacts and field inhomogeneities.
2.4. Image analysis
Before automatic preprocessing, the images were checked for artifacts andmanually aligned to the AC-PC line. Data were processed and analyzed usingSPM8 (http://www.fil.ion.ucl.ac.uk/spm) and VBM8 toolbox (http://dbm.neuro.uni-jena.de/vbm.html), for which we used the default parameters. Thus, within a unifiedsegmentation model (Ashburner and Friston, 2005), images were corrected for bias-field inhomogeneities, registered using linear (12 parameters affine) and non-lineartransformations (warped), and tissue was clustered into GM, WM, and cerebrospinalfluid (CSF). Segments were further refined by using adaptive maximum a posteri-ori estimations, which account for partial volume effects, and by applying a hidden
d prefrontal gray matter reductions in cocaine dependent individuals.
Markov random field model, as implemented in VBM8. Importantly, to preservelocal GM/WM values, we multiplied the segments by the Jacobian determinantsof the deformation field to create modulated images. Segments were smoothed byan 8 mm full-width at half-maximum (FWHM) using an isotropic Gaussian kernel.Afterwards, we conducted analysis on modulated GM and WM segments.

ARTICLE IN PRESSG Model
DAD-4380; No. of Pages 7
L. Moreno-López et al. / Drug and Alcohol Dependence xxx (2012) xxx– xxx 3
Table 1Descriptive information about patterns of drug use in cocaine-dependent individuals.
Substance Ever used (%) Unit Amount per month Duration of use Abstinence
Cocaine 100 # grams 18.92 ± 29.46 4.05 ± 3.07 36.76 ± 22.60Cannabis 76.31 # joints 109.90 ± 111.40 2.35 ± 3.17 147.45 ± 203.66MDMA 50 # tablets 9.29 ± 10.12 2.27 ± 1.90 227.68 ± 225.78Alcohol 100 # SAU 107.58 ± 114.03 7.38 ± 5.77 33.11 ± 21.77Tobacco 86.8 # cigarettes 488.48 ± 307.40 9.19 ± 7 7.8 ± 15.82
Note: Mean ± standard deviation of the mean. Duration of use is expressed in years. Abstinence duration is expressed in number of weeks.
Table 2Descriptive scores – means and standard deviations (in parentheses) of cocaine users and non-drug using controls in the five UPPS-P dimensions.
UPPS-P dimensions Cocaine-dependent individuals Non-drug using controls Test
Negative urgency 31.45 (6.79) 23.16 (6.99) (t46 = 5.14, p = 0.000)Positive urgency 32.91 (8.79) 21.66 (7.63) (t46 = 5.86, p = 0.000)
2(i(tttos
2ntcacwvfg
2etUPic(
3
3
aceaC[Sb
3
ts(p
Lack of premeditation 23.83 (5.95)
Lack of perseverance 21.26 (4.42)
Sensation seeking 30.63 (8.73)
.4.1. Global effects of patterns of drug use. Total gray and white matter volumesTGMV, TWMV) were computed by adding up the voxel values of GM/WM segments,ncluding cerebellum and sub-cortical structures. Next, the total brain volumeTBV = TGMV + TWMV) was used for computing the TGMV/TBV ratios, which servedo evaluate the global effects of patterns of drug use on brain tissue volumes usinghe backward step-wise procedure in the general linear model, including as predic-ors age, years of education, cocaine amount per month during regular use, durationf cocaine use, and age of onset of cocaine use. t-Tests were used to estimate theignificance of the regression parameters. Significance threshold was set at p < 0.05.
.4.2. Regional GM and WM differences between cocaine-dependent individuals andon-drug using controls. The general linear model implemented in SPM8 was usedo conduct voxel-wise comparisons between cocaine-dependent individuals andontrols, and to determine the relationship between consume severity factors (i.e.mount, duration and age of onset) and regional GM and WM volume variations inocaine-dependent individuals. In both analyses the total volume of GM, or WM,as modeled as a linear confound in order to account for residual global volume
ariability. The significance threshold was set at p < 0.05 after family-wise correctionor multiple comparisons (pFWE < 0.05). Significant peaks from the t-test maps areiven in MNI space.
.4.3. Regional relationships between GM and WM and measures of impulsivity andstimates of drug use. We performed SPM8 multiple regression analysis, in which weested the significances of the Group (cocaine-dependent individuals vs. controls) xPPS-P dimensions interactions across the entire brain. We only included the UPPS-
dimensions showing significant differences between the groups. The differencesn the regression coefficients as a function of group were tested using a signifi-ance threshold of p < 0.05 after family-wise correction for multiple comparisonspFWE < 0.05).
. Results
.1. Participants’ characteristics
Both groups were matched on gender, ethnicity and language,nd had statistically equivalent distributions for age. Controls,ompared to cocaine-dependent individuals, had more years ofducation [(M = 17.58, SD = 4.56) vs. (M = 12.03, SD = 3.62)]; butnalysis including education as a covariate did not alter the results.ocaine-dependent individuals had greater scores for depression(M = 5.08, SD = 3.98) vs. (M = 1.32, SD = 2.15)] and anxiety [(M = 7.05,D = 6.40) vs. (M = 1.29, SD = 2.29)], but on average both groups wereelow cut-offs for clinical significance.
.2. Trait impulsivity
Cocaine-dependent individuals had higher scores than con-
Please cite this article in press as: Moreno-López, L., et al., Trait impulsivity anDrug Alcohol Depend. (2012), doi:10.1016/j.drugalcdep.2012.02.012
rols across the five dimensions of the UPPS-P Scale. t-Testshowed these differences were significant for lack of premeditationt = 2.46, p = 0.016, Cohen’s d = 0.6), lack of perseverance (t = 2.56,
= 0.013, Cohen’s d = 0.6), negative urgency (t = 5.14, p = 0.000,
21 (3.68) (t46 = 2.46, p = 0.016)18.84 (3.63) (t46 = 2.56, p = 0.013)29.89 (7.37) (t46 = 0,34, p = 0.73)
Cohen’s d = 1.2), and positive urgency (t = 5.86, p = 0.000, Cohen’sd = 1.4). We found no significant differences for sensation seeking,such that this dimension was not further used in the subsequentanalysis.
3.3. Imaging analysis
3.3.1. Global effects. For cocaine-dependent individuals, theTGMV/TBV ratio variability was accounted for by monthly cocaineintake during regular use, t(34) = 2.19, p = 0.035, and age of onset,t(34) = −2.33, p = 0.026; the effect of duration of cocaine use alsoshowed a trend to significance, t(34) = −1.80, p = 0.080. Thesevariables explained about a 32% of the variability in TGMV/TBVratios, F(3,34) = 5.21, p = 0.005. Noteworthy, higher amounts ofcocaine (r = 0.41, p = 0.01) and younger age of onset of cocaine use(r = −.34, p = 0.04) were associated with larger TGMV/TBV. The firstresult is associated with a significant decrease in WM volumes(TWMV-amount of cocaine: r = 0.43, p = 0.006), but the secondshowed a trend in relation to reductions in GM (TGMV-age of onsetof cocaine abuse: r = −0.18, NS).
3.3.2. Regional GM and WM differences between cocaine-dependentindividuals and non-drug using controls. The cocaine group hadsignificantly lower GM and WM volumes than controls. Cocaine-dependent individuals showed lower GM volumes in a numberof sections of the OFC (left superior and medial frontal gyrus),right inferior frontal gyrus, right insula, left amygdala and parahip-pocampal gyrus, left inferior and middle temporal gyrus, andbilateral caudate (Table 3 and Fig. 1). Likewise, cocaine dependent-individuals showed lower WM volumes in the left inferior andmedial frontal gyrus, superior temporal gyrus, right anterior cin-gulate cortex, insula and caudate (Table 4 and Fig. 2).
3.3.3. Regional relationships between GM and WM and measuresof impulsivity and estimates of drug use. We found a number ofregions in which GM volumes showed differential associations withthe impulsivity dimensions of lack of premeditation and negativeurgency; in all cases, there was a significant Group x Impulsiv-ity interaction (see Table 5). The associations between impulsivitylevels and GM volumes were significantly different for cocaine-dependent individuals vs. controls in the left inferior frontal gyrus,right insula and left putamen (in relation to lack of premeditation),
d prefrontal gray matter reductions in cocaine dependent individuals.
and left middle frontal gyrus and sub-gyral (in relation to negativeurgency). Left inferior frontal gyrus was positively associated withlack of premeditation in cocaine users, whereas it was negativelycorrelated with this dimension in controls. Conversely, right insula

ARTICLE IN PRESSG Model
DAD-4380; No. of Pages 7
4 L. Moreno-López et al. / Drug and Alcohol Dependence xxx (2012) xxx– xxx
Table 3Summary of the results obtained by the gray matter voxel-wise analysis of volume reductions in cocaine-dependent individuals and non-drug using controls.
Anatomical region k Anatomical labels for peaks BA T p x y z
FrontalLobe
3342 L Medial Frontal Gyrus 11, 10 5.81 0.004 −9 27 −13Cingulate Gyrus 32, 25, 24 5.75 0.005
Frontal Lobe 2802 R Inferior Frontal Gyrus 47, 11, 34 6.39 0.000 28 21 −15Caudate R Caudate Head 5.53 0.010 9 19 −15Caudate 1118 L Caudate Head 5.52 0.010 6 −1 3Limbic Lobe/Parahippocampal Gyrus 972 L Amygdala/Hippocampus 6.42 0.000 −33 −7 −19Sub-Lobar 581 R Insula 13 5.81 0.004 52 −19 23Temporal Lobe 3095 L Middle/Inferior Temporal Gyrus 21, 22, 20 5.98 0.002 −56 −14 −18
Fig. 1. Brain regions showing significant gray matter decreases in cocaine-dependent individuals with respect to non-drug using controls.
Table 4Summary of the results obtained by the white matter voxel-wise analysis of volume reductions in cocaine-dependent individuals and non-drug using controls.
Anatomical Region k Anatomical Labels for peaks T p x y z
FrontalLobe
1181L Inferior Frontal Gyrus
6.07 0.001 −26 18 −17Sub-GyralFrontal Lobe 623 L Medial Frontal Gyrus 5.46 0.003 −20 57 −3Temporal Lobe 389 L Superior Temporal Gyrus 5.24 0.007 −48 −25 −3FrontalLobe
1651R Anterior Cingulate
6.81 0.001 7 33 −15R Medial Frontal Gyrus
Sub-Lobar 1531Caudate
5.86 0.001 12 12 −3Extra-NuclearLentiform Nucleus
TemporalLobe
674Insula
5.82 0.001 46 −26 12R Superior Temporal GyrusR Transverse Temporal Gyrus
Fig. 2. Brain regions showing significant white matter decreases in cocaine-dependent individuals with respect to non-drug using controls.
Table 5Regression slopes showing different associations between impulsivity and gray matter volumes between cocaine-dependent individuals (CDI) and non-drug-using controls(NDC) (pFWE = 0.05).
Anatomical region k Anatomical Label BA p (ˇ) Slopes Volume x y z
Lack of premeditation
Sub-lobar179 R Insula BA 47/BA
130.02 CDI−/NDC+ CDI < NDC 30 10 −3
358 L Putamen 0.05 CDI−/NDC+ CDI < NDC −21 10 −4Fontal Lobe 249 L Inferior Frontal Gyrus BA 10 0.005 CDI+/NDC− CDI < NDC −37 41 1
Please cite this article in press as: Moreno-López, L., et al., Trait impulsivity anDrug Alcohol Depend. (2012), doi:10.1016/j.drugalcdep.2012.02.012
Negative urgencyFrontalLobe
228 L Middle Frontal Gyrus BA10/46
381 R Sub-Gyral BA 8
d prefrontal gray matter reductions in cocaine dependent individuals.
0.038 CDI+/NDC− CDI < NDC −39 37 190.024 CDI+/NDC− CDI < NDC 25 24 36

ING Model
D
Alcoho
aiisamza
ag
mc
4
parriiiowmuccwiiamotta
uitt(Macreumst2Gpq
ttfism
ARTICLEAD-4380; No. of Pages 7
L. Moreno-López et al. / Drug and
nd left putamen were negatively correlated with lack of premed-tation in cocaine users, but did not correlate with this dimensionn controls. With regard to negative urgency, the sub-gyral showedignificant positive correlations in the cocaine group, whereas thessociation was negative in controls. Finally, in the case of the leftiddle frontal gyrus, none of the slopes significantly differed from
ero, but the trend was positive in cocaine-dependent individuals,nd negative in controls.
We did not find significant associations between WM volumesnd the different dimensions of impulsivity in the cocaine or controlroups.
Finally, we did not find significant associations between theain patterns of drug use (amount, duration and age of onset of
ocaine abuse) and GM and WM volumes.
. Discussion
Our results showed that cocaine-dependent individuals, com-ared to controls, have lower GM volumes in the OFC cortex,nterior cingulate, inferior frontal gyrus, insula, amygdala, tempo-al gyrus, and caudate. We also found WM reductions in adjacentegions of the anterior cingulate, inferior/middle frontal gyrus,nsula and putamen. As expected, we found significant differencesn the way that impulsivity dimensions correlates with GM volumesn cocaine users vs. controls: cocaine patients had elevated levelsf trait impulsivity, and these scores were differentially associatedith GM volumes in the left inferior frontal gyrus, insula and puta-en (lack of premeditation), and left middle frontal gyrus (negative
rgency). Inferior and medial superior frontal clusters positivelyorrelated with impulsivity in cocaine users, and negatively inontrols; conversely, insula and putamen correlated negativelyith impulsivity in cocaine patients and showed no correlations
n controls. These results point to a distinctive link between traitmpulsivity and regional GM among cocaine-dependent individu-ls, which specifically relates to frontostriatal systems regions. Theeasures of patterns of cocaine use correlated with general indices
f GM/TBV ratios but had no significant influence on GM volumes athe stricter voxel level. Overall, our findings are in agreement withhe notion that GM reductions in cocaine-dependent individualsre at least partly mediated by trait impulsivity.
Our findings of GM reductions in cocaine-dependent individ-als are largely consistent with previous neuroimaging studies
n revealing structural abnormalities in brain regions relevanto reinforcement learning (amygdala, OFC) and inhibitory con-rol (anterior cingulate, inferior frontal gyrus, insula and caudate)Ersche et al., 2011; Franklin et al., 2002; Makris et al., 2004;
atochik et al., 2003). Recent findings from fMRI connectivitynalyses also provide support to the notion that neural networksonnecting the medial frontal cortex with paralimbic and tempo-al regions are significantly less functional in cocaine users (Gut al., 2010). Biological abnormalities in these regions are thought tonderlie psychological alterations leading to overestimation of theotivational relevance of drug-related reinforcement (the impul-
ive system) and to breakdown of inhibitory processes necessaryo achieve long-term abstinence (the reflective system) (Bechara,005; Goldstein and Volkow, 2002; Redish et al., 2008; Verdejo-arcía and Bechara, 2009). These alterations are proposed to anchorervasive drug taking in the face of increasing aversive conse-uences, one of the key features of addiction (DSM-IV).
A key unresolved issue is that if these structural abnormali-ies are reflective of cocaine induced brain attrition, or are related
Please cite this article in press as: Moreno-López, L., et al., Trait impulsivity anDrug Alcohol Depend. (2012), doi:10.1016/j.drugalcdep.2012.02.012
o personality traits, or reflect a combination of both factors. Ourndings, in agreement with growing neuroscience evidence, areupportive of the impact of personality traits. However, these traitsay interact with cocaine use in producing GM alterations, as
PRESSl Dependence xxx (2012) xxx– xxx 5
suggested by the global TGMV analysis. Recent studies indicatethat, in healthy individuals, impulsivity is negatively correlatedwith midbrain and striatal D2/D3 receptors availability (Buckholtzet al., 2010; Lee et al., 2009), but this association is strongerin methamphetamine dependent individuals (Lee et al., 2009)and correlates with amphetamine-induced dopamine release inthe striatum (Buckholtz et al., 2010). Striatal D2 availability isassociated with metabolic rate in the prefrontal cortex of psychos-timulant users (Volkow et al., 2001). Therefore, different strandsof evidence indicate that frontostriatal systems mediate impul-sive behavior and thereby promote psychostimulant addiction.Nonetheless, we acknowledge that these issues can only be testedby longitudinal imaging studies in individuals at high risk of devel-oping psychostimulant dependence, or by gene association imagingstudies (Kreek et al., 2005; Verdejo-García et al., 2008).
In accordance with the notion that impulsivity is a multifacetedconstruct (Smith et al., 2007; Whiteside and Lynam, 2001), ourresults showed differential links between impulsivity dimensionsand GM volumes. Lack of premeditation was positively associatedwith left inferior frontal gyrus GM in cocaine users but not con-trols. This region is importantly involved in the representationof expected values (Bermpohl et al., 2010); such that persistentstimulation by drug-expected rewards could have induced GMenlargements linked to an increased tendency to rapidly engagein non-adequately forethought behavior. On the other hand, bothinsula and putamen have been involved in drug-related neuroadap-tations related to attentional bias and craving (Childress et al., 2008;Luijten et al., 2011; Volkow et al., 2006), such that reduced volumesmay predict increased prepotency of drug-related action tenden-cies. On the other hand, negative urgency was positively associatedwith GM in subgyral BA 8 in cocaine users, negatively in controls.This region has been previously associated with the experience ofuncertainty (Volz et al., 2003), which is a key aspect of the associa-tive learning processes mediating expected drug effects (D’Souzaand Duvauchelle, 2008). In fact, adjacent regions of the medialfrontal gyrus (BA 10) display increased activation during uncertainoutcome selection in psychostimulant users (Leland et al., 2006).Therefore, in this case the rationale could be similar to that positedfor inferior frontal gyrus; repeated exposure to uncertainty-loadedscenarios may be associated with increased subgyral volumesand higher risk to trigger impulsive acts when under emotionallyuneasy conditions. Remarkably, in both dimensions (lack of pre-meditation and urgency) there was a positive correlation betweentrait impulsivity and GM volume in the inferior/middle frontalgyrus of cocaine-dependent individuals, a pattern directly opposedto the association in controls. Given that GM in these regions isreduced in cocaine users, one may speculatively argue that highimpulsivity may render some cocaine users more similar to con-trols. Nonetheless, given the role of these regions in emotionalvaluation, greater GM in these areas may hold specific emotionalmotives in cocaine users: high impulsive individuals, with greaterGM in inferior/middle frontal gyrus, might be – for example – highlysensitive to craving (Garavan et al., 2000). More research is war-ranted to resolve this issue.
Strengths of this study include the adequate sample size, thesound clinical characterization and the agreement between resultsfrom the different analytical approaches (between-group differ-ences and regressions testing the link between impulsivity andGM). On the other hand, we should also acknowledge a numberof limitations. The cross-sectional design precludes us from draw-ing conclusions about the causality of GM deficits; one way outof addressing this issue would be by using longitudinal designs,
d prefrontal gray matter reductions in cocaine dependent individuals.
but these studies are costly and convey important ethical con-cerns and methodological complexities. Furthermore, we cannotcompletely rule out the influence of depression and anxiety lev-els, although they were on average below the cut-off for clinical

ING Model
D
6 Alcoho
sepuccdct
R
MsaiifSQ
C
ihaEt
C
prysp
R
A
AB
B
B
B
B
C
C
D
ARTICLEAD-4380; No. of Pages 7
L. Moreno-López et al. / Drug and
ignificance and they had very low impact on regression mod-ls when included. Finally, although we discuss our results asertaining to cocaine dependence, polysubstance abuse is almostbiquitous among cocaine-dependent individuals. However, theurrent sample was carefully selected for relative specificity ofocaine related problems and low use of other drugs. The illegalrugs more frequently and intensely co-abused in the sample wereannabis and MDMA, but estimates of the use of these drugs failedo predict regional GM.
ole of funding source
This study has been funded by grants SEJ2006-08278, Spanishinistry of Education and Science and P07-HUM-03089, Andalu-
ia Council of Science and Innovation (PI: Miguel Pérez-García)nd grant COPERNICO, Plan Nacional sobre Drogas: Spanish Min-stry of Health (PI: Antonio Verdejo-García). Laura Moreno-Lópezs supported by a FPU predoctoral research grant (AP2007-03583)rom the Spanish Ministry of Science and Innovation. Emmanuel A.tamatakis is supported by the Stephen Erskine Fellowship fromueens’ College, Cambridge.
ontributors
Laura Moreno-López and Antonio Verdejo-García have workedn the conception and design of the manuscript. Andrés Catenaas carried out the analysis and interpretation of the data as wells the revision of the manuscript. María José Fernández-Serrano,lena Delgado-Rico and Miguel Pérez-García have contributed tohe revision of the manuscript.
onflict of interest
The authors declare that, except for income received from theirrimary employer, no financial support or compensation has beeneceived from any individual or corporate entity over the past threeears for research or professional service and there are no per-onal financial holdings that could be perceived as constituting aotential conflict of interest.
eferences
lia-Klein, N., Parvaz, M.A., Woicik, P.A., Konova, A.B., Maloney, T., Shumay, E.,Wang, R., Telang, F., Giegon, Al., Wang, G.J., Fowler, J.S., Tomasi, D., Volkow, N.D.,Goldstein, R.Z., 2011. Gene × disease interaction on orbitofrontal gray matter incocaine addiction. Arch. Gen. Psychiatry 68, 283–294.
shburner, J., Friston, K.J., 2005. Unified segmentation. Neuroimage 26, 839–851.arrós-Loscertales, A., Garavan, H., Bustamante, J.C., Ventura-Campos, N., Llopis, J.J.,
Belloch, V., Parcet, M.A., Avila, C., 2011. Reduced striatal volume in cocaine-dependent patients. Neuroimage 56, 1021–1026.
echara, A., 2005. Decision making, impulse control and loss of willpower to resistdrugs: a neurocognitive perspective. Nat. Neurosci. 8, 1458–1463.
elin, D., Mar, A.C., Dalley, J.W., Robbins, T.W., Everitt, B.J., 2008. Highimpulsivity predicts the switch to compulsive cocaine-taking. Science 320,1352–1355.
ermpohl, F., Kahnt, T., Dalanay, U., Hägele, C., Sajonz, B., Wegner, T., Stoy, M., Adli, M.,Kruger, S., Wrase, J., Strohle, A., Bauer, M., Heinz, A., 2010. Altered representationof expected value in the orbitofrontal cortex in mania. Hum. Brain Mapp. 31,958–969.
uckholtz, J.W., Treadway, M.T., Cowan, R.L., Woodward, N.D., Li, R., Ansari, M.S.,Baldwin, R.M., Schwartzman, A.N., Shelby, E.S., Smith, C.E., Kessler, R.M., Zaid,D.H., 2010. Dopaminergic network differences in human impulsivity. Science329, 532.
hildress, A.R., Ehrman, R.N., Wang, Z., Li, Y., Sciortino, N., Hakun, J., Jens, W., Suh, J.,Listerud, J., Marquez, K., Franklin, T., Langleben, D., Detre, J., O’Brien, C.P., 2008.Prelude to passion: limbic activation by “unseen” drug and sexual cues. PLoSONE 3, e1506.
yders, M.A., Smith, G.T., Spillane, N.S., Fischer, S., Annus, A.M., Peterson, C.,
Please cite this article in press as: Moreno-López, L., et al., Trait impulsivity anDrug Alcohol Depend. (2012), doi:10.1016/j.drugalcdep.2012.02.012
2007. Integration of impulsivity and positive mood to predict risky behavior:development and validation of a measure of positive urgency. Psychol. Assess.19, 107–118.
alley, J.W., Fryer, T.D., Brichard, L., Robinson, E.S., Theobald, D.E., Lääne, K., Pena,Y., Murphy, E.R., Shah, Y., Probst, K., Abakumova, I., Aigbirhio, F.I., Richards,
PRESSl Dependence xxx (2012) xxx– xxx
H.K., Hong, Y., Baron, J.C., Everitt, B.J., Robbins, T.W., 2007. Nucleus accumbensD2/3 receptors predict trait impulsivity and cocaine reinforcement. Science 315,1267–1270.
D’Souza, M.S., Duvauchelle, C.L., 2008. Certain or uncertain cocaine expectationsinfluence accumbens dopamine responses to self-administered cocaine andnon-rewarded operant behavior. Eur. Neuropsychopharmacol. 18, 628–638.
EMCDDA European Monitoring Centre for Drugs and Drug Addiction, 2009. AnnualReport 2009: The State of the Drugs Problem in Europe. Publications Office ofthe European Union, Luxembourg.
Ersche, K.D., Barnes, A., Simon Jones, P., Morein-Zamir, S., Robbins, T.W., Bullmore,E.T., 2011. Abnormal structure of frontostriatal brain systems is associated withaspects of impulsivity and compulsivity in cocaine dependence. Brain 134,2013–2024.
Ersche, K.D., Bullmore, E.T., Craig, K.J., Shabbir, S.S., Abbott, S., Müller, U., Ooi, C., Suck-ling, J., Barnes, A., Sahakian, B.J., Merlo-Pich, E.V., Robbins, T.W., 2010. Influenceof compulsivity of drug abuse on dopaminergic modulation of attentional biasin stimulant dependence. Arch. Gen. Psychiatry 67, 632–644.
Franklin, T.R., Acton, P.D., Maldjian, J.A., Gray, J.D., Croft, J.R., Dackis, C.A., O’Brien,C.P., Childress, A.R., 2002. Decreased gray matter concentration in the insular,orbitofrontal, cingulate, and temporal cortices of cocaine patients. Biol. Psychi-atry 51, 134–142.
Garavan, H., Pankiewicz, J., Bloom, A., Cho, J.K., Sperry, L., Ross, T.J., Salmeron, B.J.,Risinger, R., Kelley, D., Stein, E.A., 2000. Cue-induced cocaine craving: neu-roanatomical specificity for drug users and drug stimuli. Am. J. Psychiatry 157,1789–1798.
Goldstein, R.Z., Volkow, N.D., 2002. Drug addiction and its underlying neurobiologi-cal basis: neuroimaging evidence for the involvement of the frontal cortex. Am.J. Psychiatry 159, 1642–1652.
Gu, H., Salmeron, B.J., Ross, T.J., Geng, X., Zhan, W., Stein, E.A., Yang, Y., 2010. Meso-corticolimbic circuits are impaired in chronic cocaine users as demonstrated byresting-state functional connectivity. Neuroimage 53, 593–601.
Koob, G.F., Volkow, N.D., 2010. Neurocircuitry of addiction. Neuropsychopharma-cology 35, 217–238.
Kreek, M.J., Bart, G., Lilly, C., LaForge, K.S., Nielsen, D.A., 2005. Pharmacogenetics andhuman molecular genetics of opiate and cocaine addictions and their treatments.Pharmacol. Rev. 57, 1–26.
Lee, B., London, E.D., Poldrack, R.A., Farahi, J., Nacca, A., Monterosso, J.R., Mumford,J.A., Bokarius, A.V., Dahlborn, M., Mukherjee, J., Bilder, R.M., Brody, A.L., Man-delkern, M.A., 2009. Striatal dopamine d2/d3 receptor availability is reducedin methamphetamine dependence and is linked to impulsivity. J. Neurosci. 29,14734–14740.
Leland, D.S., Arce, E., Feinstein, J.S., Paulus, M.P., 2006. Young adult stimulant users’increased striatal activation during uncertainty is related to impulsivity. Neu-roimage 33, 725–731.
Luijten, M., Veltman, D.J., van den Brink, W., Hester, R., Field, M., Smits, M., Franken,I.H., 2011. Neurobiological substrate of smoking-related attentional bias. Neu-roimage 54, 2374–2381.
Makris, N., Gasic, G.P., Seidman, L.J., Goldstein, J.M., Gastfriend, D.R., Elman, I.,Albaugh, M.D., Hodge, S.M., Ziegler, D.A., Sheahan, F.S., Caviness Jr., V.S., Tsuang,M.T., Kennedy, D.N., Hyman, S.E., Rosen, B.R., Breiter, H.C., 2004. Decreased abso-lute amygdala volume in cocaine addicts. Neuron 44, 729–740.
Matochik, J.A., London, E.D., Eldreth, D.A., Cadet, J.L., Bolla, K.I., 2003. Frontal corticaltissue composition in abstinent cocaine abusers: a magnetic resonance imagingstudy. Neuroimage 19, 1095–1102.
Matsuo, K., Nicoletti, M., Nemoto, K., Hatch, J.P., Peluso, M.A., Nery, F.G., Soares, J.C.,2009. A voxel-based morphometry study of frontal gray matter correlates ofimpulsivity. Hum. Brain Mapp. 30, 1188–1195.
Narayana, P.A., Datta, S., Tao, G., Steinberg, J.L., Moeller, F.G., 2010. Effect of cocaine onstructural changes in brain: MRI volumetry using tensor-based morphometry.Drug Alcohol Depend. 111, 191–199.
Redish, A.D., Jensen, S., Johnson, A., 2008. A unified framework for addiction: vul-nerabilities in the decision process. Behav. Brain Sci. 31, 415–437.
Sim, M.E., Lyoo, I.K., Streeter, C.C., Covell, J., Sarid-Segal, O., Ciraulo, D.A., Kim, M.J.,Kaufman, M.J., Yurgelun-Todd, D.A., Renshaw, P.F., 2007. Cerebellar gray mattervolume correlates with duration of cocaine use in cocaine-dependent subjects.Neuropsychopharmacology 32, 2229–2237.
Smith, G.T., Fischer, S., Cyders, M.A., Annus, A.M., Spillane, N.S., 2007. On the valid-ity and utility of discriminating among impulsivity-like traits. Assessment 14,155–170.
Substance Abuse and Mental Health Services Administration, 2010. Results from the2009 National Survey on Drug Use and Health: Mental Health Findings (Officeof Applied Studies, NSDUH Series H-39, HHS Publication No. SMA 10-4609),Rockville, MD.
Tanabe, J., Tregellas, J.R., Dalwani, M., Thompson, L., Owens, E., Crowley, T., et al.,2009. Medial orbitofrontal cortex gray matter is reduced in abstinent substance-dependent individuals. Biol. Psychiatry 65, 160–164.
Verdejo-García, A., Bechara, A., 2009. A somatic marker theory of addiction. Neu-ropharmacology 56, 48–62.
Verdejo-García, A., Lawrence, A.J., Clark, L., 2008. Impulsivity as a vulnerabilitymarker for substance-use disorders: review of findings from high-risk research,problem gamblers and genetic association studies. Neurosci. Biobehav. Rev. 32,
d prefrontal gray matter reductions in cocaine dependent individuals.
777–810.Verdejo-García, A., Lozano, O., Moya, M., Alcázar, M.A., Pérez-García, M., 2010. Psy-
chometric properties of a Spanish version of the UPPS-P impulsive behaviorscale: reliability, validity and association with trait and cognitive impulsivity. J.Pers. Assess. 92, 1532–7752.

ING Model
D
Alcoho
V
V
V
ARTICLEAD-4380; No. of Pages 7
L. Moreno-López et al. / Drug and
erdejo-García, A.J., López-Torrecillas, F., Aguilar de Arcos, F., Pérez-García, M., 2005.Differential effects of MDMA, cocaine, and cannabis use severity on distinc-tive components of the executive functions in polysubstance users: a multipleregression analysis. Addict. Behav. 30, 89–101.
Please cite this article in press as: Moreno-López, L., et al., Trait impulsivity anDrug Alcohol Depend. (2012), doi:10.1016/j.drugalcdep.2012.02.012
olkow, N.D., Wang, G.J., Telang, F., Fowler, J.S., Logan, J., Childress, A.R., Jayne, M., Ma,Y., Wong, C., 2006. Cocaine cues and dopamine in dorsal striatum: mechanismof craving in cocaine addiction. J. Neurosci. 26, 6583–6588.
olkow, N.D., Chang, L., Wang, G.J., Fowler, J.S., Ding, Y.S., Sedler, M., Logan, J.,Franceschi, D., Gatley, J., Hitzemann, R., Gifford, A., Wong, C., Pappas, N., 2001.
PRESSl Dependence xxx (2012) xxx– xxx 7
Low level of brain dopamine D2 receptors in methamphetamine abusers: asso-ciation with metabolism in the orbitofrontal cortex. Am. J. Psychiatry 158,2015–2021.
Volz, K.G., Schubotz, R.I., von Cramon, D.Y., 2003. Predicting events of
d prefrontal gray matter reductions in cocaine dependent individuals.
varying probability: uncertainty investigated by fMRI. Neuroimage 19,271–280.
Whiteside, S.P., Lynam, D.R., 2001. The five factor model and impulsivity: using astructural model of personality to understand impulsivity. Pers. Individ. Dif. 30,669–689.

[ANEXO II]

Neural Correlates of the Severity of Cocaine, Heroin, Alcohol, MDMA and Cannabis
Use in Polysubstance Abusers: A Resting-PET Brain Metabolism Study
Laura Moreno-Lópeza, Emmanuel A. Stamatakisb, Maria José Fernández-Serranoc, Manuel
Gómez-Ríod, Antonio Rodríguez-Fernándezd, Miguel Pérez-Garcíaa,e,f and Antonio
Verdejo-Garcíaa,f
a Department of Personality, Evaluation and Psychological Treatment, University of
Granada, Campus de Cartuja s/n, 18071, Spain.
b Division of Anaesthesia, School of Clinical Medicine, University of Cambridge, U.K.
c Departament of Psychology, University of Jaén, Campus Las Lagunillas s/n, 23071, Spain
d Service of Nuclear Medicine of the Hospital Virgen de las Nieves of Granada, Avda. de
las Fuerzas Armadas 2, 18014, Spain.
e Centro de Investigación Biomédica en Red de Salud Mental, CIBERSAM. University of
Granada, Spain.
f Institute of Neurosciences Federico Olóriz, University of Granada, Avda. de Madrid, s/n,
Facultad de Medicina, 18013, Spain.
Correspondence should be sent to:
Laura Moreno-López
Departamento de Personalidad, Evaluación y Tratamiento Psicológico.
Universidad de Granada. Campus de Cartuja s/n, 18071, Granada, Spain.
Phone: +34 958 242 948; Fax: +34 958 243 749;
E-mail: [email protected]
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65

Abstract
Introduction: Functional imaging studies of addiction following protracted
abstinence have not been systematically conducted to look at the associations between
severity of use of different drugs and brain dysfunction. Findings from such studies may be
relevant to implement specific interventions for treatment. The aim of this study was to
examine the association between resting-state regional brain metabolism (measured with
18F-fluorodeoxyglucose Positron Emission Tomography (FDG-PET) and the severity of
use of cocaine, heroin, alcohol, MDMA and cannabis in a sample of polysubstance users
with prolonged abstinence from all drugs used. Methods: Our sample consisted of 49
polysubstance users enrolled in residential treatment. We conducted correlation analyses
between estimates of use of cocaine, heroin, alcohol, MDMA and cannabis and brain
metabolism (BM) (using Statistical Parametric Mapping voxel-based (VB) whole-brain
analyses). In all correlation analyses conducted for each of the drugs we controlled for the
co-abuse of the other drugs used. Results: The analysis showed significant negative
correlations between severity of heroin, alcohol, MDMA and cannabis use and BM in the
dorsolateral prefrontal cortex (DLPFC) and temporal cortex. Alcohol use was further
associated with lower metabolism in frontal premotor cortex and putamen, and stimulants
use with parietal cortex. Conclusions: Duration of use of different drugs negatively
correlated with overlapping regions in the DLPFC, whereas severity of cocaine, heroin and
alcohol use selectively impact parietal, temporal, and frontal-premotor/basal ganglia regions
respectively. The knowledge of these associations could be useful in the clinical practice
since different brain alterations have been associated with different patterns of execution
that may affect to the rehabilitation of these patients.
Key words: Cocaine, Heroin, Alcohol, MDMA, Cannabis, Drug severity, Positron
Emission Tomography.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65

1. Introduction
Drug addiction has been associated with neuroadaptations in brain systems involved
in motivation, memory and executive control [1]. Functional neuroimaging studies have
revealed that the use of different classes of drugs is associated with dysfunctions in a range
of overlapping brain regions including ventromedial and dorsolateral prefrontal cortex
(DLPFC), anterior cingulate cortex, inferior frontal gyrus, insula, amygdala, basal ganglia
and cerebellum [2]. These dysfunctions may explain the overlapping cognitive deficits
observed in users of different drugs, including working memory, inhibitory control,
flexibility or decision-making deficits [3]. In addition, some specific effects have been
described for the association between cocaine use and the parietal cortex [4,5,6], heroin use
and the temporal cortex [7], MDMA use and occipital, hippocampus and thalamic regions
[8,9], and cannabis use and the premotor cortex [10,11]. These associations are congruent
with relatively specific neuropsychological deficits pertaining to attention and cognitive
control in cocaine users, long-term memory in opiate users, visuospatial memory in MDMA
users, and psychomotor function in cannabis users [3,7]. In spite of the available evidence,
in vivo studies have not been systematically conducted to look at associations between
severity of use of different drugs and brain dysfunction. In fact, several neuroimaging
studies have failed to detect such link or have provided controversial results (reviews in 12).
These negative findings and controversies might be linked to the impact of relevant
confounding variables. One of these is the fact that neuroimaging studies have frequently
tested drug users currently using drugs or having brief periods of abstinence (24-48 h) [13,
14]; under these conditions, the presumed link between lifetime drug use and brain
dysfunction could be masked by several other factors, including recent drug use or
psychological symptoms associated with withdrawal and short-term abstinence [2]. Another
key variable for consideration is the concurrent use of multiple types of substances, which
can introduce significant confounds in the interpretation of the data.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65

The current study aimed to specifically address the association between lifetime
estimates of use of different types of drugs and brain functioning (as measured by 18F-
fluorodeoxyglucose Positron Emission Tomography (FDG-PET), statistically controlling
for the concurrent use of other drugs in a sample of polysubstance users with prolonged
abstinence from all drugs. Specifically, we examined the link between lifetime estimates of
amount and duration of use of cocaine, heroin, alcohol, MDMA and cannabis and brain
metabolism (BM). Based on previous findings we hypothesized that exposure to all
substances would be significantly associated with overlapping reductions of metabolism in
frontal, limbic, striatal and cerebellar regions, whereas cocaine and heroin use would
specifically correlate with parietal and temporal cortices respectively.
2. Methods
2.1 Participants
Forty-nine substance dependent individuals (SDI), forty-one men and eight women
with a mean age of 32.67 years, were recruited during their stay in an inpatient treatment
program at the centre �Proyecto Hombre� in Granada (Spain). This centre provides a
controlled setting for the treatment of substance use disorders. Selection criteria for
participants in this study were (i) meeting the DSM-IV criteria for substance dependence;
(ii) absence of documented comorbid mood or personality disorders, as assessed by clinical
reports; (iii) absence of documented head injury or neurological disorders; and (iv) to have
minimum abstinence duration from all drugs consumed (except for tobacco) of 15 days
before testing. In fact, SDI participants had overall much longer abstinence duration (mean
of 32.94, SD=11.25 weeks); for that reason it was possible to rule out the presence of
withdrawal symptoms or brain function alterations associated with the acute or short term
effects of the drugs. None of the participants were experiencing withdrawal symptoms as
assessed by routine medical examination or were enrolled in opioid substitution treatments
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65

with methadone or other pharmacological treatments (e.g., benzodiazepines) during the
course of the assessment interview/PET evaluations.
2.2 Testing protocols and procedures
This study was approved by the Human Subjects Committee of the University of
Granada. One-step Syva urine drug screens for amphetamines, benzodiazepines, cannabis,
cocaine and opiates were conducted to confirm abstinence at time of testing. Participants
were evaluated individually in two different sessions. During the first session, the
participants were provided with information about the study and were encouraged to ask
questions to ascertain they fully understood the rational for the study and were clear on the
details for participation. The participants then provided written consent and were assessed
with the Interview for Research on Addictive Behaviours [15] in the �Proyecto Hombre�
installations�. During the second session, each patient was accompanied by a therapist to
the nuclear medicine service of the �Virgen de las Nieves� hospital, where the PET
neuroimaging session took place. This second session usually took place one week after the
first session was completed.
2.3 Tools
2.3.1 Drug Use Information
The Interview for Research on Addictive Behaviour [15] was used to examine the
severity of drug use. This interview evaluates, by means of a brief interview, the quantity
(average dosing), frequency (number of drug taking episodes per month), and duration
(years of duration) of the use of different substances that can produce physical or
psychological dependence, including cocaine, heroin, alcohol, MDMA and cannabis which
were the main drugs of choice in the present study. For every substance the participant had
actually used, the following information was requested: (i) the average dose of each target
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65

drug taken in each episode of use (number of grams for cocaine and heroin, number of units
for alcohol, considering that 25 cl. of hard liquor (e.g., scotch) equals 2 units, while 25 cl. of
wine or beer equals 1 unit, number of pills for MDMA and number of cigarettes for
cannabis); (ii) the frequency of these consumption episodes per month (daily, between one
and three times upon a week, once a week, between one and three times upon a month, or
once a month); and (iii) the number of years that elapsed since the onset of use. From this
information, we obtained two indices for each of the drugs used: (i) Amount (average dose x
frequency), an index of monthly use of the drug; and (ii) Duration of drug use, measuring
number of years of exposure to the drug. The main sociodemographic and clinical features
from the sample are displayed in Table 1.
- Please insert Table 1 here-
2.3.2 PET Image Acquisition
PET scans were acquired with an ECAT/931 scanner (Siemens CTI ECAT
EXACT), at the service of nuclear medicine centre of the hospital �Virgen de las Nieves� in
Granada (Spain). For each individual, we obtained 20 minute emission scans 30 minutes
after the injection of one dose of 200 MBq of FDG administrated only after levels of
glycemia had been checked (they must be below 120 mg/dl) [16]. The subjects were
scanned with their eyes open and ears unplugged in a dimly illuminated quiet room. Raw
data were processed using an iterative reconstruction technique (OSEM method: 10
iterations, 32 subsets). Images were reoriented in transaxial, coronal and saggital planes.
For the analyses reported here the reconstructed images used had 47 axial slices each with
an in-plane resolution of 2.57x2.57 and a slice thickness of 3.38mm.
2.4 Data analysis
2.4.1 Preprocessing of PET images
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65

PET images were converted from DICOM to NIFTI format and were spatially
normalized to the SPM PET template (Wellcome Department of Cognitive Neurology,
London, UK) using linear (12 affine) and non-linear normalization. An average image of all
the normalized images was obtained and this acted as a study specific PET template. In this
manner we constructed a template in the same modality, from the same scanner as the
images we wanted to normalize. The raw PET images were then spatially normalized with
linear affine and nonlinear parameters to this study-specific template. Visual inspection
revealed this optimized spatial normalization technique produced satisfactory spatial
normalization for the PET image of every volunteer.
2.4.2 Statistical Analysis
The spatially normalized PET images were smoothed with an 8mm3 isotropic
Gaussian filter and were statistically modeled using the General Linear Model in SPM5.
Linear regressions were used to carry out several analyses in order to identify brain areas
with a significant correlation between uptake values and measures of the amount and
duration of cocaine, heroin, alcohol, MDMA and cannabis use. Not all subjects used all the
drugs but since virtually all participants had regularly abused two or more substances, the
analyses were conducted on the whole group and results were interpreted in relation to
polysubstance use. In each of the analysis, we controlled for the effects of the other
substances concurrently used (e.g., analysis of severity of cocaine use controlled for the
effects of co-abuse of heroin, alcohol, MDMA and cannabis), age and years of education.
We report local maximum peaks that survive a voxel threshold of p<0.001 (uncorrected for
multiple comparisons, cluster size �100 voxels). We chose to report our findings at this
threshold based on numerous similar studies of PET imaging in drug addiction [17,18].
3. Results
The main results are summarized in Table 2.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65

3.1 Amount of use
Voxel-based analyses showed negative correlations between lifetime amount of
cocaine, alcohol and cannabis use and several brain regions. The amount of cocaine used
was negatively correlated with a cluster encompassing the left inferior parietal lobule
extending to the left postcentral gyrus (Figure 1.1). The amount of alcohol used was
negatively associated with three clusters: one cluster included the left middle and superior
temporal cortex; a second cluster included the bilateral superior frontal cortex extending to
the left DLPFC and the right supplementary motor area; and a third cluster included the
right DLPFC extending to the superior frontal gyrus (Figure 1.2). The amount of cannabis
use showed a negative correlation with two clusters: one cluster included the left inferior
frontal gyrus (pars triangularis) extending to the DLPFC, and a second cluster included the
right DLPFC extending to the superior frontal cortex (Figure 1.3). We did not find
significant correlations between the amount of heroin or MDMA use and BM measures.
3.2 Duration of use
Voxel-based analyses showed negative correlations between duration of heroin,
alcohol and MDMA use and several brain regions. Duration of heroin use showed a
negative correlation with three clusters, which encompassed the right inferior, middle and
superior temporal cortex extending to the right supramarginal and inferior frontal gyrus
(Figure 1.4). Duration of alcohol use was negatively correlated with four clusters. One
cluster included the right inferior temporal cortex extending to the middle aspect of this
area. A second cluster included the right putamen and pallidum. A third cluster
encompassed the right precentral and postcentral gyrus and the fourth cluster included the
right fusiform and parahippocampal gyrus (Figure 1.5). Finally, we found negative
correlations between the duration of MDMA use and BM in three clusters: the first cluster
included the left postcentral and inferior parietal gyrus; the second cluster included the right
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65

inferior frontal gyrus (pars triangularis) extending to the DLFPC; and the third cluster
included the right superior temporal pole extending to the middle part of this region (Figure
1.6).
� Please insert Table 2 here �
4. Discussion
Our findings showed that the estimates of severity of use of heroin, alcohol, MDMA
and cannabis were negatively associated with DLPFC and temporal cortex regional
metabolism in polysubstance dependent individuals with prolonged abstinence from all
these drugs. Alcohol use was further associated with lower metabolism in frontal premotor
cortex and putamen. Furthermore, severity of stimulants use (cocaine and MDMA) was
uniquely associated with inferior parietal/postcentral cortex metabolism. These associations
were established after a prolonged period of abstinence, and after controlling for multiple
confounding variables, thus surpassing the limitations of previous functional neuroimaging
studies that were not specifically designed to study prolonged abstinence, and were
confounded by important variables, such as withdrawal related symptoms or concurrent use
of multiple substances. The associations found are in agreement with our initial predictions,
and with the neuropsychological deficits typically aligned with the chronic use of these
substances; these include reduced competency in executive functions (related to all classes
of drugs), episodic memory (mainly observed on heroin, alcohol, MDMA and cannabis
users), motor control (alcohol and cannabis users), and visuospatial attention (stimulants
users) (see meta-analyses in 3,8,19,20).
As expected, the estimates of amount or duration of different classes of drugs were
negatively associated with overlapping regions within the DLPFC (BA 9, 45, 46). This
region has been linked with several of the key neuroadaptations associated with drug
addiction, including drug conditioning, loss of self-control, and stimulus-driven compulsive
behavior [2]. This finding is also in agreement with results from several neuropsychological
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65

studies showing cognitive deficits on working memory, planning or inhibition processes,
which are associated with DLPFC functioning, across users of several drugs [3]. We also
found negative associations between estimates of duration of heroin, alcohol, and MDMA
use and regional metabolism on partially overlapping sections of the temporal cortex. These
associates may substantiate the links between the intensity and duration of involvement
with these drugs and the degree of memory attrition, which has been observed in
longitudinal studies with users of these substances [21,22]. It is worth mentioning that
duration of heroin use was shorter to that of cocaine and alcohol use; however, previous
brain structural findings indicate that duration of heroin use is a critical factor leading to
brain damage, even in users with periods of use below five years [23]. Surprisingly, we
failed to find significant associations between the severity of the substances used and
overlapping limbic, striatal and cerebellar regions but this result could be explained from
the fact that these regions play a greater role during current use and short-term abstinence
[24]. In addition, we found relatively specific negative correlations between amount of
cocaine use and regional metabolism in the inferior parietal cortex, and between alcohol
duration and regions involved in motor programming and control (frontal precentral and
putamen). The link between cocaine use and parietal dysfunction has been observed on
previous functional imaging studies during cognitive challenges of attention and executive
control [6,25,26]. The link between alcohol use and precentral and basal ganglia
dysfunction is in agreement with structural imaging findings showing that function of these
regions is significantly reduced in alcohol users compared to other forms of addiction [27].
Additional evidence comes from volumetric studies which associated volumetric measures
of these regions and deficient motor skills in alcoholics [28]. These more specific findings
could be used to design tailored interventions aimed to address specific aspects of frontal-
subcortical executive functions in different profiles of substance abusers.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65

An important implication of this study is the fact that the link between severity of
drug use and BM is still observable in substance abusers with prolonged periods of
abstinence (an average of eight months in this sample). Therefore, quantity and duration of
drug use may still be affect brain functioning after several months of successful cessation of
drug use. This is particularly relevant in view of recent findings indicating that certain
patterns of brain dysfunction can reliably predict mid-term and long-term relapse.
Permanent brain alterations may also have important repercussions for the rehabilitation of
SDI, since these alterations can affect the ability of SDI to assimilate the contents and
participate in the activities of the rehabilitation programs [29,30]. In fact, preliminary
evidence suggests that cognitive rehabilitation techniques, such as errorless learning may be
successful in compensating chronic cognitive deficits in substance abusers [31]. Thus, the
brain networks impacted by severity of drug abuse can play a central role in clinical
outcome and relapse and should become priority targets for therapeutic interventions.
Several possible limitations of our study should be considered and addressed by
future research. Our findings were not obtained in �pure� users of each of the substances
but in mostly polysubstance users. Although this limitation is inherent to the clinical
literature on addiction (i.e., �pure� users of one single drug are quite rare among substance
abusers enrolled in treatment programs), we attempted to control for the co-abuse of other
drugs using statistics. A related confounding variable was the use of nicotine, which is
ubiquitous among drug users but was not specifically assessed or controlled for.
Furthermore, due to the use of a correlational design, this study cannot yield conclusions
about cause-effect relationships between the use of drug and resting BM. However, given
the agreement of our results with both resting state PET, activation PET/fMRI and addiction
neuropsychology studies, we could hypothesize that, at least in part, the alterations observed
in this population could be due to the quantity and duration related effects of the
consumption of the substances under consideration in this study.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65

One last point to consider is that our findings could be due to premorbid brain
alterations or the results of the interaction between the premorbid alterations and the
neurotoxic effects of drug use. This question should be addressed through longitudinal
designs. A related limitation is that no comparison with a healthy control database was
performed. As we argued before, a healthy control group was not needed to test the main
predictions of our study; however, the lack of controls has stopped us from making any
assumption about the direction of these findings in relation to the healthy population.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65

References
1. Koob GF, Volkow ND (2010) Neurocircuitry of addiction. Neuropsychopharmacology
35: 217-238.
2. Goldstein RZ, Volkow ND (2011) Dysfunction of the prefrontal cortex in addiction:
neuroimaging findings and clinical implications. Nat Rev Neurosci 12: 652-669.
3. Fernández-Serrano MJ, Pérez-García M, Verdejo-García A (2011) What are the specific
vs. generalized effects of drugs of abuse on neuropsychological performance? Neurosci
Biobehav Rev 35: 377-406.
4. Aron JL, Paulus MP (2007) Location, location: using functional magnetic resonance
imaging to pinpoint brain differences relevant to stimulant use. Addiction 1: 33-43.
5. Lane SD, Steinberg JL, Ma L, Hasan KM, Kramer LA, et al. (2010) Diffusion tensor
imaging and decision making in cocaine dependence. PLoS One 5: e11591.
6. Tomasi D, Goldstein RZ, Telang F, Maloney T, Alia-Klein N, et al. (2007) Widespread
disruption in brain activation patterns to a working memory task during cocaine abstinence.
Brain Res 1171: 83-92.
7. Gruber SA, Silveri MM, Yurgelun-Todd DA (2007) Neuropsychological consequences
of opiate use. Neuropsychol Rev 17: 299-315.
8. Kish SJ, Lerch J, Furukawa Y, Tong J, McCluskey T, et al. (2010) Decreased cerebral
cortical serotonin transporter binding in ecstasy users: a positron emission
tomography/[(11)C]DASB and structural brain imaging study. Brain 133: 1779-1797.
9. de Win MM, Jager G, Booij J, Reneman L, Schilt T, et al. (2008) Neurotoxic effects of
ecstasy on the thalamus. Br J Psychiatry 193: 289-296.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65

10. King GR, Ernst T, Deng W, Stenger A, Gonzales RM, et al. (2011) Altered brain
activation during visuomotor integration in chronic active cannabis users: relationship to
cortisol levels. J Neurosci 31: 17923-17931.
11. Pillay SS, Rogowska J, Kanayama G, Gruber S, Simpson N, et al. (2008) Cannabis and
motor function: fMRI changes following 28 days of discontinuation. Exp Clin
Psychopharmacol 16: 22-32.
12. Martín-Santos R, Fagundo AB, Crippa JA, Atakan Z, Bhattacharyya S, et al. (2010)
Neuroimaging in cannabis use: a systematic review of the literature. Psychol Med 40: 383-
98.
13. Goldstein RZ, Alia-Klein N, Tomasi D, Carrillo JH, Maloney T, et al. (2009) Anterior
cingulate cortex hypoactivations to an emotionally salient task in cocaine addiction. Proc
Natl Acad Sci U S A 106: 9453-9458.
14. Zubieta JK, Heitzeg MM, Xu Y, Koeppe RA, Ni L, et al. (2005) Regional cerebral
blood flow responses to smoking in tobacco smokers after overnight abstinence. Am J
Psychiatry 162: 567-577.
15. Verdejo-García AJ, López-Torrecillas F, Aguilar de Arcos F, Pérez-García M (2005)
Differential effects of MDMA, cocaine, and cannabis use severity on distinctive
components of the executive functions in polysubstance users: a multiple regression
analysis. Addict Behav 30: 89-101.
16. Boellaard R, O�Doherty MJ, Weber WA, Mottaghy FM, Lonsdale MN, et al. (2010)
FDG PET and PET/CT: EANM procedure guidelines for tumour PET imaging: version 1.0.
Eur J Nucl Med Mol Imaging 37: 181-200.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65

17. Fehr C, Hohmann N, Gründer G, Dielentheis TF, Buchholz HG, et al. (2007) Tiagabine
does not attenuate alcohol-induced activation of the human reward system.
Psychopharmacology (Berl) 191: 975-983.
18. Volkow ND, Fowler JS, Wang GJ, Telang F, Logan J, et al. (2010) Cognitive control of
drug craving inhibits brain reward regions in cocaine abusers. Neuroimage 49: 2536-2543.
19. Jovanovski D, Erb S, Zakzanis KK (2005) Neurocognitive deficits in cocaine users: a
quantitative review of the evidence. J Clin Exp Neuropsychol 27: 189-204.
20. Zakzanis KK, Campbell Z, Jovanovski D (2007) The neuropsychology of ecstasy
(MDMA) use: a quantitative review. Hum Psychopharmacol 22: 427-435.
21. Hanson KL, Cummins K, Tapert SF, Brown SA (2011) Changes in neuropsychological
functioning over 10 years following adolescent substance abuse treatment. Psychol Addict
Behav 25: 127-142
22. de Sola S, Tarancón T, Peña-Casanova J, Espadaler JM, Langohr K, et al. (2008)
Auditory event-related potentials (P3) and cognitive performance in recreational ecstasy
polydrug users: evidence from a 12-month longitudinal study. Psychopharmacology (Berl)
200: 425-437
23. Yuan Y, Zhu Z, Shi J, Zou Z, Yuan F, et al. (2009) Gray matter density negatively
correlates with duration of heroin use in young lifetime heroin-dependent individuals. Brain
Cogn 71: 223-228.
24. Breiter HC, Rosen BR (1999) Functional magnetic resonance imaging of brain reward
circuitry in the human. Ann N Y Acad Sci 877: 523-547.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65

25. Barrós-Loscertales A, Bustamante JC, Ventura-Campos N, Llopis JJ, Parcet MA, et al.
(2011) Lower activation in the right frontoparietal network during a counting Stroop task in
a cocaine-dependent group. Psychiatry Res 194: 111-118.
26. Bustamante JC, Barrós-Loscertales A, Ventura-Campos N, Sanjuán A, Llopis JJ, et al.
(2011) Right parietal hypoactivation in a cocaine-dependent group during a verbal working
memory task. Brain Res 1375: 111-119.
27. van Holst RJ, de Ruiter MB, van den Brink W, Veltman DJ, Goudriaan AE (2012) A
voxel-based morphometry study comparing problem gamblers, alcohol abusers, and healthy
controls. Drug Alcohol Depend. In press.
28. Sullivan EV, Deshmukh A, De Rosa E, Rosenbloom MJ, Pfefferbaum A (2005) Striatal
and forebrain nuclei volumes: contribution to motor function and working memory deficits
in alcoholism. Biol Psychiatry 57: 768-776.
29. Johnson-Greene D, Adams KM, Gilman S, Junck L (2002) Relationship between
neuropsychological and emotional functioning in severe chronic alcoholism. Clin
Neuropsychol 16: 300-309.
30. Zinn S, Stein R, Swartzwelder HS (2004) Executive functioning early in abstinence
from alcohol. Alcohol Clin Exp Res 28: 1338-1346.
31. Pitel AL, Perruchet P, Vabret F, Desgranges B, Eustache F, et al. (2009) The advantage
of errorless learning for the acquisition of new concepts' labels in alcoholics. Psychol Med
40: 497-502.
Tzourio-Mazoyer N, Landeau B, Papathanassiou D, Crivello F, Etard O, et al. (2002)
Automated anatomical labeling of activations in SPM using a macroscopic anatomical
parcellation of the MNI MRI single-subject brain. Neuroimage 15: 273-289.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65

Figure Legends
Figure 1. Areas of reduced metabolism predicted by the amount or duration of drug
use: (1) amount of cocaine, (2) amount of alcohol, (3) amount of cannabis (4) duration of
heroin (5) duration of alcohol and (6) duration of MDMA. Areas of reduced metabolism are
superimposed on a T1 weighted MRI image in MRICRON. MNI coordinates are shown
underneath each panel.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65

Tables
Table 1 Sociodemographic data and clinical features of the participants
Subjects Characteristics
Gender
Male 41
Female 8
Mean S.D. Maximum Minimum
Age (years) 32.67 8.03 53 21
Education (years) 9.73 2.34 17 5
Amount of drug usea
Substance Mean S.D. Maximum Minimum
Cocaine (g) (46) 48.69 44.83 180 0
Heroin (g) (16) 9.12 20.63 120 0
Alcohol (units) (45) 571.24 489.34 1800 0
MDMA (pills) (23) 13.41 24.31 120 0
Cannabis (cigarettes) (37) 148.57 190.64 750 0
Duration of drug use (years)
Substance Mean S.D. Maximum Minimum
Cocaine 7.95 5.95 23 0
Heroin 1.76 3.69 17 0
Alcohol 10.85 7.67 27 0
MDMA 1.40 2.28 8 0
Cannabis 8.31 8.13 29 0
Abstinence (weeks) 32.94 11.25 60 12
a Amount of monthly use of the drug calculated using the average dose (per episode) ×
frequency (per month).
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65

Table 2 Relation between severity of drug use and PET uptake.
Substanc Voxels-Based BA KE T MNI Amount
Cocaine Parietal Inf L 40 176 3.98 -58 -38 54
Alcohol Temporal Mid L 21 267 4.04 -66 -2 -10
Frontal Sup L 9 308 3.96 -20 30 36
Frontal Sup R 6 365 3.73 22 2 60
Frontal Mid R 46 372 3.82 26 22 34
Cannabis
Frontal Inf Tri L 45 355 4.40 -44 22 30
Frontal Mid R 46 125 4.02 24 48 28
Duration
Heroin Frontal Mid R 46 876 3.77 44 36 34
Temporal Inf R 20 257 4.58 62 -16 -32
Temporal Sup R 22 1090 4.19 66 -44 20
Alcohol Temporal Inf R 20 238 4.22 64 -16 -30
Putamen R 472 3.71 20 -2 8
Precentral R 6 274 3.67 50 2 34
Fusiform R 20 299 3.65 30 -6 -40
MDMA Postcentral L 40 131 3.73 -28 -40 48
Frontal Inf Tri R 45 139 3.67 44 34 20
Temporal Pole Sup R 20 561 3.67 32 12 -30
Voxel-based analyses at p<0.001 (uncorrected, cluster size > 100 voxels). Labels were
obtained using the aal toolbox (Tzourio-Mazoyer et al., 2002). Broadmann areas were
obtained using MRICron (Rorden and Brett, 2000). KE indicate the number of voxels
included in the cluster. Stereotaxic coordinates are those listed in SPM5. Inf, inferior;
Sup, superior; L, left; R, right; Supp, supplementary; Mid, middle; Orb, orbitalis.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65


[ANEXO III]

1Q2
2
3
4Q1
5
678910
11
1213141516171819202122232425
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
Psychiatry Research: Neuroimaging xxx (2012) xxx–xxx
PSYN-09914; No of Pages 8
Contents lists available at SciVerse ScienceDirect
Psychiatry Research: Neuroimaging
j ourna l homepage: www.e lsev ie r .com/ locate /psychresns
RO
OF
Neural correlates of hot and cold executive functions in polysubstance addiction:Association between neuropsychological performance and resting-PETbrain metabolism
Laura Moreno-López a,⁎, Emmanuel Andreas Stamatakis b, María José Fernández-Serrano a,Manuel Gómez-Río c, Antonio Rodríguez-Fernández c, Miguel Pérez-García a,d,e, Antonio Verdejo-García a,d
a Department of Personality, Evaluation and Psychological Treatment, University of Granada, Spainb Division of Anaesthesia, School of Clinical Medicine, University of Cambridge, UKc Service of Nuclear Medicine of the Hospital Virgen de las Nieves of Granada, Spaind Institute of Neurosciences Federico Olóriz, University of Granada, Spaine Centro de Investigacion Biomédica en Red de Salud Mental, CIBERSAM, University of Granada, Spain
⁎ Corresponding author at: Departamento de Persmiento Psicológico, Universidad de, Granada, Campus dSpain. Tel.: +34 958 242 948; fax: +34 958 243 749.
E-mail address: [email protected] (L. Moreno-López).
0925-4927/$ – see front matter © 2012 Elsevier Irelanddoi:10.1016/j.pscychresns.2012.01.006
Please cite this article as: Moreno-López, L.ation between neuropsychological perform
Pa b s t r a c t
a r t i c l e i n f o26
27
28
29
30
31
32
33
34
35
Article history:Received 7 April 2010Received in revised form 20 October 2011Accepted 16 January 2012Available online xxxx
Keywords:Brain substratesDecision-makingExecutive functioningPositron emission tomography
36
37
38
39
40
41
RECTEDThe study of substance abuse related neuropsychological deficits and brain alterations may provide a better
understanding of the neuroadaptations associated with addiction. In this study we investigated the associa-tion between performance on neuropsychological tests of cold and hot executive functions and regional brainmetabolism (measured with Positron Emission Tomography – PET) in a sample of 49 substance dependentindividuals (SDI). Neuropsychological performance in the SDI group was compared to that of a non-drugusing control group of 30 participants, and associated with two sets of PET-derived dependent measures:one based on regions of interest (examining mean uptake on selected regions), and a second based onvoxel uptake measures (using Statistical Parametric Mapping voxel-based whole-brain analyses). Behavioralanalyses showed that SDI had poorer performance than controls across executive function and emotion pro-cessing measures. Regression models showed that SDI's performance in cold executive tests was associatedwith regional metabolism in the dorsolateral prefrontal cortex (DLPFC), mid-superior frontal gyrus, superiorand inferior temporal gyrus and inferior parietal cortex, whereas performance in hot executive functions wasassociated with DLPFC, mid-superior frontal gyrus, anterior and mid-posterior cingulate, and temporal andfusiform gyrus. These results are discussed in terms of their relevance for the understanding of cognitive dys-function and neuroadaptations linked to addiction.
© 2012 Elsevier Ireland Ltd. All rights reserved.
4243
R59
60
61
62
63
64
65
66
67
68
69
70
71
72
UNCO1. Introduction
Contemporary neurobiological models consider addiction to be abrain disorder that involves long-term neuroadaptations leading topersistent drug use despite increasing negative consequences(Goldstein and Volkow, 2002; Verdejo-García et al., 2004; Baler andVolkow, 2006). These neuroadaptations are thought to affect neuralsystems involved in motivation, emotion, learning, memory and ex-ecutive functioning (Ersche et al., 2008; Verdejo-García and Bechara,2009). From this perspective, it is crucial to understand both the long-term neuropsychological consequences of drug abuse and their neu-ral substrates.
There is compelling evidence that substance dependent individ-uals (SDI) present widespread deficits in neuropsychological
73
74
75
76
77
onalidad, Evaluación y Trata-e Cartuja S/N, 18071, Granada,
Ltd. All rights reserved.
, et al., Neural correlates of hance..., Psychiatry Research:
functioning, and that these deficits are especially prominent in so-called executive functions (Fernández-Serrano et al., 2011). Executivefunctions are defined as a set of higher-order abilities involved in theemployment, monitoring, and regulation of goal-directed behaviors.Recent evidence suggests that these functions can be classified intotwo broad domains: (i) cold executive functions, involved in the pro-cessing of relatively abstract, context-free information, which arelinked to more dorsal and lateral regions of the prefrontal cortex,and (ii) hot executive functions, which come to play when emotion-ally laden information is concerned, and are linked to the functioningof the orbitofrontal cortex (OFC) (Kerr and Zelazo, 2004).
Although executive dysfunction in SDI has been extensively de-scribed, the study of the neurobiological substrates of executive dys-function in the context of drug addiction is still a growing researcharea (Goldstein and Volkow, 2002; Lundqvist, 2010). A number of func-tional magnetic resonance imaging (fMRI) studies have shown signifi-cant associations between performance on cognitive tasks indexingcold aspects of executive functions and abnormal activation of brainsystems in SDI. Polysubstance abusers have shown abnormally
ot and cold executive functions in polysubstance addiction: Associ-Neuroimaging (2012), doi:10.1016/j.pscychresns.2012.01.006

TE
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125126127128129130131132133134135136137138139140141142143144
145146147148149150151152153154155156157
158
159160161162163164165166167
168
169170171172173174175
176177178179180181182183184185186187188189
190191192193194195196197198199
Table 1 t1:1
Sociodemographic data and clinical features of the participants.t1:2t1:3SDI (n=49) HC (n=30)
t1:4Variable N % N %
t1:5Gendert1:6Male 41 84 24 80t1:7Female 8 16 6 20t1:8Mean S.D. Mean S.D.t1:9Age (years) 32.67* 8.03 26.4* 8.03t1:10Education (years) 9.73* 2.34 11.63* 2.04
* p≤0.001. t1:11
2 L. Moreno-López et al. / Psychiatry Research: Neuroimaging xxx (2012) xxx–xxx
UNCO
RREC
increased or decreased activation of the dorsolateral prefrontal cortex(DLPFC), orbitofrontal cortex (OFC), temporo-parietal regions and thecerebellum during performance on working memory (Desmond et al.,2003; Jager et al., 2006; Tomasi et al., 2007), response inhibition (Dao-Castellana et al., 1998; Bolla et al., 2004; Eldreth et al., 2004; Formanet al., 2004; Lee et al., 2005), and cognitive flexibility tasks (Gilman etal., 1990; Goldstein et al., 2004). Nevertheless, very few studies have in-vestigated the neural correlates of indices of hot executive functions inSDI. These abilities have been associatedwith the OFC and the cingulatecortex in cognitive neuroscience studies (Anderson et al., 2006;Heberlein et al., 2008; Hartstra et al., 2010), and they have been foundto be dysfunctional in polysubstance abusers even aftermid-term absti-nence (circa sixmonths) (Fernández-Serrano et al., 2011). For example,two neuropsychological studies have revealed deficits in goal-drivenself-regulation in psychostimulant users using the Revised Strategy Ap-plication Test (R-SAT) (Halpern et al., 2004; Verdejo-García et al., 2007),but no studies to date have investigated the neural substrates of self-regulation in addiction. Another key factor for addiction initiation andmaintenance is adaptive decision-making, which is typically indexedusing the Iowa Gambling Task (IGT) (Bechara et al., 1994). Performanceon this task has been associatedwith hyperperfusion in the anterior cin-gulate cortex (ACC), middle and superior frontal gyrus and DLPFC insmall samples of shortly abstinent cocaine-dependent individuals(Adinoff et al., 2003; Tucker et al., 2004). However, because of the keyrole of decision-making in mid- and long-term drug relapse (Paulus etal., 2005; Passetti et al., 2008), there is a need to extend these findingsin larger samples ofmid-term abstinent SDI. Similarly, the neural corre-lates of emotional processing and perception must be considered inlight of their association with hot aspects of executive functions in SDI(Verdejo-García et al., 2007), and their predictive role in drug relapse(Lubman et al., 2009). Only two previous neuroimaging studies have fo-cused on this domain, and they have found decreased ACC and amygda-la activations in alcohol users exposed to facial emotional expressions(Salloum et al., 2007; Marinkovic et al., 2009).
In this study we aimed to investigate the neuropsychological per-formance of a sample of SDI and healthy controls (HC) on tests tap-ping on cold (i.e., updating, inhibition and flexibility) and hot (i.e.,self-regulation, decision-making and emotion perception) aspects ofexecutive functions, and the association between the neuropsycho-logical performance of SDI on those tests and PET-indexed regionalmetabolism. We hypothesize that neuropsychological performancewill be poorer in SDI than HC and that it will be associated withDLPFC, ACC and temporal regions for tests of cold executive functions,and with OFC, ACC and limbic regions for tests of hot executivefunction.
2. Methods
2.1. Participants
Forty-nine SDI were recruited from an inpatient treatment program at the centre“Proyecto Hombre” in Granada (Spain). The treatment has a mean duration of 6 monthsand integrates key elements of cognitive behavioral psychology and systemic approachesto provide complete social rehabilitation and integration. Selection criteria for participantsin this studywere (1) meeting the DSM-IV criteria for substance dependence at the onsetof the treatment; (2) absence of documented comorbid mood or personality disorders, asassessed by clinical reports; (3) absence of documented head injury or neurological disor-ders, as assessed by clinical reports and PET images; (4) to be literate enough to performany literacy tests, as assessed by the TAP reading skills test (Del Ser et al., 1997) and (5) tohave aminimum abstinence duration of 15 days before testing. In fact, the SDI included inthe sample had overall much longer abstinence duration (M=32.94, S.D.=11.25 weeks),allowing us to rule out acute or short term effects of drug intake. Since virtually all partic-ipants had regularly abused two or more substances, the analyses were conducted on thewhole group and results were interpreted in relation to polysubstance use. Urine toxicol-ogy screening (one-step Syva rapid tests) for the different drugs used were conductedroutinely in the SDI, sowe can rule out the use of these substances during the entire periodof abstinence. None of the participants were experiencing withdrawal symptoms beforeor during neuropsychological/PET testing as assessed by routine medical examination.Likewise, none of the participants were enrolled in opioid substitution treatments withmethadone or other pharmacological treatments during the course of the
Please cite this article as: Moreno-López, L., et al., Neural correlates of hation between neuropsychological performance..., Psychiatry Research:
neuropsychological/PET testing. A group of drug-free healthy individuals was recruitedas a comparison group for neuropsychological performance, but these participants werenot PET-scanned. HC were recruited through local advertisements and snowball commu-nication among adult people from the community. Selection criteria for these groupwere:(1) absence of current or past substance abuse, excluding past cannabis use and past orcurrent smoking or social drinking (in cases of current social drinking, exclusion criteriaincluded acute use of alcohol during the last 24 h before evaluation), (2) absence of docu-mentedmajor psychiatric disorders, (3) absence of documented head injury or neurolog-ical disorder and (4) not being on any medication that could affect the normalneuropsychological functioning. This group only had exposure to alcohol use, having aregular mean use of 8.85 standard units per week (S.D.=20.7) and a mean duration ofuse of 6.7 years (S.D.=7.3). The main sociodemographic and clinical features from thesamples are displayed in Table 1.
OO
F
2.2. Testing protocols and procedures
This study was approved by the Human Subjects Committee of the University ofGranada. All participants signed an informed consent form before participating in thestudy. The testing protocol, including the neuropsychological assessment and the PETscanning were administered into two sessions that lasted approximately 70 mineach. The control group only participated in the first session. During the first session,participants completed the Interview for Research on Addictive Behaviors and per-formed the neuropsychological tests protocol. During the second session (up to7 days later), SDI were scanned at the nuclear medicine service of the “Virgen de lasNieves” hospital.
D P
R2.3. Instruments
2.3.1. Patterns of drug useThe information about the severity of drug addiction was collected by the Interview
for Research on Addictive Behavior (IRAB; López-Torrecillas et al., 2001) (see supple-mentary material). This is a brief interview that collects information about the quantity(average dose), frequency (consumption episodes by month), and duration (years ofduration) of the use of a series of drugs of abuse, including cocaine, heroin, and alcohol.The main drug use information of the SDI group is displayed in Table 2.
2.3.2. Neuropsychological testsWe administered a battery of tests designed to assess several domains related to
cold and hot executive functions, including working memory, response inhibition, cog-nitive flexibility, self-regulation, decision-making and emotion processing.
The tests used were Letter Number Sequencing (LNS) (Wechsler Adult IntelligenceScale – WAIS-III, Wechsler, 1997), Stroop (Golden, 1978), Category Test (DeFilippis,2002), Revised Strategy Application Test (R-SAT) (Levine et al., 2000), Iowa GamblingTask (IGT) (Bechara et al., 1994) and Ekman Faces Test (EFT) (Young et al., 2002). Alltests were administered in a fixed order and according to standard instructions. Thedescription of the instruments is provided in the supplementary material.
Due to technical problems during data acquisition, we could not obtain the perfor-mance indices for some of the tests in the SDI group. Therefore, the number of SDI par-ticipants included in the analyses of LNS was 40, 39 for Stroop, 47 for Category Test, 49for R-SAT and IGT, and 48 for EFT.
2.3.3. PET image acquisitionPET scans were acquired with the ECAT/931 scanner (Siemens CTI ECAT EXACT), at
the nuclear medicine facility of the “Virgen de las Nieves” hospital in Granada, (Spain).For each individual, we obtained 20 minute emission scans 30 min after injection ofFDG resting in quiet conditions. The subjects were scanned with their eyes open andears unplugged in a dimly illuminated quiet room. Raw data were processed usingan iterative reconstruction (OSEM method: 10 iterations, 32 subsets). Images werereoriented in transaxial, coronal and saggital planes. For the analyses reported herethe reconstructed images used had 47 axial slices each with an in-plane resolution of2.57×2.57 and a slice thickness of 3.38 mm.
ot and cold executive functions in polysubstance addiction: Associ-Neuroimaging (2012), doi:10.1016/j.pscychresns.2012.01.006

T200
201202203204205206207208209210211
212213214215216
217
218219220221222223
224225226227228229230
231232233234235236237238239240241242243244245246
247248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
271
272
273
274
275
276
277
278
279
280
281
282
283
284
285
286
Table 2t2:1
Amount and duration of drug use in the substance dependence individuals.t2:2t2:3 Quantity of use
t2:4 Substance Averagea S.D. Maximum Minimum
t2:5 Cocaine (46) 48.69 44.83 180 0t2:6 Cannabis (37) 148.57 190.64 750 0t2:7 Heroin (16) 9.12 20.63 120 0t2:8 MDMA (18) 13.41 24.31 120 0t2:9 Alcohol (45) 571.24 489.34 1800 0
t2:10 Duration of use (years)
t2:11 Substance Duration S.D. Maximum Minimum Age ofonset
t2:12 Cocaine 7.95 5.95 23 0 23.26t2:13 Cannabis 8.31 8.13 29 0 20.91t2:14 Heroin 1.76 3.69 17 0 24.75t2:15 MDMA 1.40 2.28 8 0 23.66t2:16 Alcohol 10.85 7.67 27 0 20.87t2:17 Abstinenceb 32.94 11.25 60 12
a Average dose (per episode)×Frequency (per month). A dose is units in alcohol,grams in cocaine and heroin, fags in cannabis and pills in MDMA.t2:18
b Abstinence in weeks. The number in brackets is the number of substance depen-dence individuals that consumed the substance.t2:19
Table 3 t3:1
Descriptive scores on the different indices of executive functioning.t3:2t3:3Variable Group p-valuea F-value d-value
t3:4SDI HC
t3:5LNSb 9.75±2.27 15.13±2.33 0.000* 32.12 −2.34t3:6Stroopc −1.76±5.76 4.15±7.24 0.000* 7 −0.92t3:7Category testd 65.67±26 31.80±25.33 0.000* 11.48 1.32t3:8R-SATe 83.78±17.14 93.98±5.71 0.013* 3.83 −0.73t3:9IGTf −1.98±21.06 37.20±26.16 0.000* 17.90 −1.70t3:10Ekman faces testg 47.50±4.49 53±4.61 0.000* 10.03 −1.21
a pb0.05. t3:11b Letter and number sequence total score. t3:12c Stroop interference score. t3:13d Category test total errors. t3:14e Revised strategy application test percentage brief items. t3:15f Iowa gambling test total score. t3:16g Ekman faces test total identifications. t3:17
3L. Moreno-López et al. / Psychiatry Research: Neuroimaging xxx (2012) xxx–xxx
UNCO
RREC
2.4. Data analysis
2.4.1. Preprocessing of PET imagesThe PET images were converted from DICOM to NIFTI format, were spatially nor-
malized to the SPM5 PET template (Wellcome Department of Cognitive Neurology,London, UK) using linear (12 affine) and non-linear normalization. An average of allthe normalized images was obtained and this acted as a study specific PET template.In this manner we constructed a template in the samemodality, from the same scanneras the images we wanted to normalize. The raw PET images were then spatially nor-malized with linear affine and nonlinear parameters to this study-specific template. Vi-sual inspection revealed this optimized spatial normalization technique producedsatisfactory spatial normalization for the PET image of every volunteer. Finally, the im-ages were smoothed with an 8 mm3 isotropic Gaussian filter.
2.4.2. ROI analysisHaving achieved satisfactory spatial normalisation we used the Marsbar tool
(http://marsbar.sourceforge.net- Brett et al., 2002) to extract 90 regions of interest(ROIs) in the entire brain. We used the AAL atlas as a reference for these ROIs(Tzourio-Mazoyer et al., 2002).
2.4.3. Statistical analyses
2.4.3.1. Behavioral analysis. ANCOVAmodels were conducted to test neuropsychologicalperformance differences between the groups while controlling for age and educationin SPSS 15.0 for Windows (SPSS Inc., Chicago IL). Cohen's d estimations of effect sizeswere calculated according to formulations proposed in (Zakzanis, 2001). The associa-tion between length of abstinence and neuropsychological performance was exploredusing bivariate correlations in SPSS 15.0.
2.4.3.2. ROI analysis. Relative glucose uptake on standardized uptake value (SUV) wereused to obtain the mean uptake values for each ROI. Next, these values were correlatedwith the indices of neuropsychological performance. In a second analysis, those ROIsthat showed significant associations with neuropsychological indices were also corre-lated with estimates of severity of drug use. In both cases, we used partial correlationsand age as confounding variable in SPSS 15.0. Results are showed at p≤0.01 andp≤0.05 respectively (uncorrected for multiple comparisons).
2.4.3.3. Voxel based analysis. Linear regressions were used to identify clusters with a sig-nificant correlation between uptake values and indices of neuropsychological perfor-mance using age as confounding factor. Scans were corrected for differences in globalactivity by including proportional scaling in the regression model. We report localmaximum peaks that survive a voxel threshold of pb0.001 (uncorrected for multiplecomparisons, cluster size>100 voxels). Although this threshold offers only modestprotection again risk of type I error, it has been chosen in multiples PET studiesunder similar assumptions (i.e. Brooks et al., 2009; Yaouhi et al., 2009). The relation-ship between duration of abstinence and resting glucose metabolism was exploredby using linear regression models but no associations survived at a threshold ofpb0.001, and therefore it was not further considered. We did not use treatment seek-ing as a variable in our analyses because all our volunteers were on treatment, andmore specifically on the same modality of treatment (i.e., therapeutic community). Fi-nally, we carried out a second analysis in which the mean uptake of the regions wefound to be associated with neuropsychological performance in whole brain voxel-based analysis were correlated with estimates of drug use using age as confounding
Please cite this article as: Moreno-López, L., et al., Neural correlates of hation between neuropsychological performance..., Psychiatry Research:
ED P
RO
OF
variable in SPSS 15.0. Results are showed at p≤0.01 and p≤0.05 respectively (uncor-rected for multiple comparisons).
3. Results
3.1. Neuropsychological performance
Results showed that SDI had significantly poorer performance thanHC on all of the neuropsychological tests administered (Table 3). Calcu-lation of effect sizes showed large effect sizes (d>0.8) for LNS (workingmemory), Stroop (response inhibition), Category Test (cognitive flexi-bility), IGT (decision-making) and EFT (emotion processing), andmedi-um effect sizes (d>0.5) for R-SAT (self-regulation) (Table 3). We onlyfound a significant negative correlation between length of abstinenceand performance on LNS (r=−0.303).
3.2. ROI analyses
3.2.1. Association between neuropsychological performance and ROIsThese results are shown in Table 4. We found significant correla-
tions between number of hits on LNS (working memory) and cerebralmetabolism (CM) at the right middle temporal pole (r=−0.367), be-tween R-SAT proportion of brief items (self-regulation) at the right cal-carine (r=0.391) and posterior cingulum bilaterally (left r=0.440)(right r=0.454), and between IGT net score (decision-making) and theright middle (r=0.390) and superior frontal cortex (r=0.376). Therewere not significant correlations between the remaining tests and theROIs at p≤0.01.
3.2.2. Association between the ROIs associated with neuropsychologicalperformance and estimates of drug use
We failed to find significant associations between measures of se-verity of drug use and cerebral metabolism at a pb0.01 threshold. Byrelaxing the alpha level to pb0.05, we only found a positive correla-tion between amount of cocaine use and brain metabolism in theright middle temporal pole (r=0.301).
3.3. Voxel-based analyses
The main results from these analyses are summarized in Table 4.The results presented in text and table correspond to an alpha levelof pb0.001 (uncorrected, cluster size>100 voxels), but for demon-stration purposes the figures display statistical parametric maps thre-sholded at uncorrected voxel level of pb0.01.
3.3.1. Cold executive functions –Whole brain voxel-based correlates in SDIFor working memory, there was a positive correlation between
LNS and one cluster with peaks at the superior temporal gyrus(51–29 3) and inferior temporal gyrus (51–72 0); this cluster
ot and cold executive functions in polysubstance addiction: Associ-Neuroimaging (2012), doi:10.1016/j.pscychresns.2012.01.006

RO
OF
287
288
289
290
291
292
293
294
295
296
Table 4t4:1
Relationship between neuropsychological performance and PET uptake using both ROI and voxel based methods.t4:2t4:3 Instrument ROIs Voxel-based analysis BA KE t p MNI Talairach
t4:4 LNS R middletemporal pole (+) R superior temporal gyrus (+) BA22 621 5.23 0.000 52 −30 2 51 −29 3t4:5 R inferior temporal gyrus (+) BA37 261 3.77 0.000 52 −74 −4 51 −72 0t4:6 L paracentral lobule (−) BA5 502 4.67 0.000 −20 −38 52 −20 −34 50t4:7 L superior frontal gyrus (−) BA8 449 3.95 0.000 −20 28 52 −20 30 46t4:8 Stroop L pallidus (−) 191 3.82 0.000 −16 −12 −4 −16 −12 −3t4:9 L middle frontal gyrus (−) BA10 160 3.72 0.000 −34 64 10 −34 62 6t4:10 Category test L superior frontal gyrus (+) BA10 127 4.25 0.000 −26 68 4 −26 66 0t4:11 R middle frontal gyrus (−) BA6 268 3.59 0.000 50 10 48 50 12 44t4:12 L inferior parietal lobule (−) BA40 198 3.74 0.000 −46 −48 38 −46 −45 37t4:13 R-SAT R calcarine (+)
R Posterior cingulate (+)L Posterior cingulate (+)
R posterior cingulate (+) BA30 5811 4.12 0.000 18 −68 10 18 −65 12t4:14 R anterior cingulate (+) BA32 702 3.75 0.000 14 32 16 14 32 13t4:15 R precentral gyrus (+) BA6 416 3.96 0.000 22 −20 64 22 −16 60t4:16 R inferior temporal gyrus (+) BA20 152 3.58 0.000 40 2 −48 40 −0 −40t4:17 L middle temporal gyrus (−) BA21 464 4.48 0.000 −56 10 −20 −55 9 −17t4:18 IGT R middle frontal (+)
R superior frontal (+)R middle frontal gyrus (+) BA6 945 4.91 0.000 26 −6 60 26 −3 55
t4:19 L middle frontal gyrus (+) BA6 377 3.71 0.000 −28 2 54 −28 4 50t4:20 R middle frontal gyrus (+) BA6 222 3.94 0.000 54 −12 56 53 14 51t4:21 L superior frontal gyrus (+) BA10 212 3.67 0.000 −18 68 14 −18 67 10t4:22 L superior frontal gyrus (+) BA8 141 3.72 0.000 −20 44 52 −20 45 46t4:23 Ekman Faces Test Lingual gyrus (−) BA18 308 4.10 0.000 0 −102 −14 0 −99 −7t4:24 Fusiform gyrus (−) BA19 191 3.90 0.000 −36 −78 −18 −36 −76 −11
L, left; R, right; (+) positive association; (−) negative association. Correlation ROI analyses are reported at p≤0.01 (uncorrected). Voxel-based analyses at pb0.001 (uncorrected,cluster size>100 voxels). Talairach coordinates were obtained by applying the mni2tal function in MATLABR2006a (http://imaging.mrc-cbu.cam.ac.uk/imaging/MniTalairach) tothe MNI coordinates obtained from SPM5. The corresponding anatomical nameswere obtained using the TalairachClient application – version 2.4.2. – (http://www.talairach.org/client.html)for peak and nearest grey matter.t4:25
4 L. Moreno-López et al. / Psychiatry Research: Neuroimaging xxx (2012) xxx–xxx
included right superior and inferior lateral temporal regions extend-ing medially. LNS negatively correlated with a cluster in the left post-central region extending to parietal cortex, precuneus andparacentral lobule (peak at paracentral lobule, −20 −34 50), and asecond cluster which encompassed regions in the left frontal cortex
UNCO
RRECTA) Letter Number sequencing negative (b
L -20 52
B) Stroop negative correlations (blue).
L -34 -16
C) Category Test negative (blue) and pos
L -45 -26
Fig. 1. Images showing the areas where cold execut
Please cite this article as: Moreno-López, L., et al., Neural correlates of hation between neuropsychological performance..., Psychiatry Research:
D Pextending to the left DLPFC (peak at superior frontal gyrus, −20 30
46) (Fig. 1 – Panel A).For response inhibition, there was a negative correlation between
the Stroop interference score and two different clusters. The first clus-ter correspond to the left thalamus, hippocampus and pallidum (peak
Elue) and positive (yellow) correlations.
R
R
itive (yellow) correlations.
45 R
ive functions correlated with brain metabolism.
ot and cold executive functions in polysubstance addiction: Associ-Neuroimaging (2012), doi:10.1016/j.pscychresns.2012.01.006
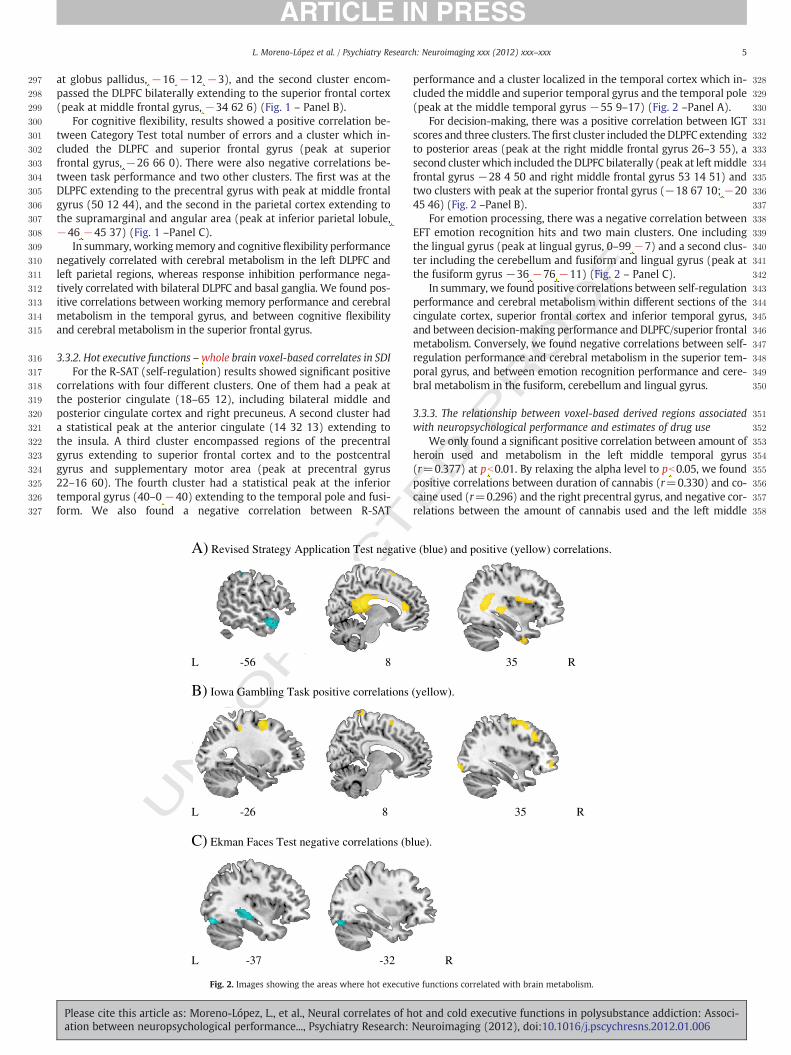
297
298
299
300
301
302
303
304
305
306
307
308
309
310
311
312
313
314
315
316
317
318
319
320
321
322
323
324
325
326
327
328
329
330
331
332
333
334
335
336
337
338
339
340
341
342
343
344
345
346
347
348
349
350
351
352
353
354
355
356
357
358
5L. Moreno-López et al. / Psychiatry Research: Neuroimaging xxx (2012) xxx–xxx
at globus pallidus, −16 −12 −3), and the second cluster encom-passed the DLPFC bilaterally extending to the superior frontal cortex(peak at middle frontal gyrus, −34 62 6) (Fig. 1 – Panel B).
For cognitive flexibility, results showed a positive correlation be-tween Category Test total number of errors and a cluster which in-cluded the DLPFC and superior frontal gyrus (peak at superiorfrontal gyrus, −26 66 0). There were also negative correlations be-tween task performance and two other clusters. The first was at theDLPFC extending to the precentral gyrus with peak at middle frontalgyrus (50 12 44), and the second in the parietal cortex extending tothe supramarginal and angular area (peak at inferior parietal lobule,−46 −45 37) (Fig. 1 –Panel C).
In summary, workingmemory and cognitive flexibility performancenegatively correlated with cerebral metabolism in the left DLPFC andleft parietal regions, whereas response inhibition performance nega-tively correlated with bilateral DLPFC and basal ganglia. We found pos-itive correlations between working memory performance and cerebralmetabolism in the temporal gyrus, and between cognitive flexibilityand cerebral metabolism in the superior frontal gyrus.
3.3.2. Hot executive functions –whole brain voxel-based correlates in SDIFor the R-SAT (self-regulation) results showed significant positive
correlations with four different clusters. One of them had a peak atthe posterior cingulate (18–65 12), including bilateral middle andposterior cingulate cortex and right precuneus. A second cluster hada statistical peak at the anterior cingulate (14 32 13) extending tothe insula. A third cluster encompassed regions of the precentralgyrus extending to superior frontal cortex and to the postcentralgyrus and supplementary motor area (peak at precentral gyrus22–16 60). The fourth cluster had a statistical peak at the inferiortemporal gyrus (40–0 −40) extending to the temporal pole and fusi-form. We also found a negative correlation between R-SAT
UNCO
RRECTA) Revised Strategy Application Test negativ
L -56 8
B) Iowa Gambling Task positive correlations
L -26 8
C) Ekman Faces Test negative correlations (b
L -37 -32
Fig. 2. Images showing the areas where hot executi
Please cite this article as: Moreno-López, L., et al., Neural correlates of hation between neuropsychological performance..., Psychiatry Research:
ED P
RO
OF
performance and a cluster localized in the temporal cortex which in-cluded the middle and superior temporal gyrus and the temporal pole(peak at the middle temporal gyrus −55 9–17) (Fig. 2 –Panel A).
For decision-making, there was a positive correlation between IGTscores and three clusters. The first cluster included theDLPFC extendingto posterior areas (peak at the right middle frontal gyrus 26–3 55), asecond cluster which included the DLPFC bilaterally (peak at leftmiddlefrontal gyrus −28 4 50 and right middle frontal gyrus 53 14 51) andtwo clusters with peak at the superior frontal gyrus (−18 67 10; −2045 46) (Fig. 2 –Panel B).
For emotion processing, there was a negative correlation betweenEFT emotion recognition hits and two main clusters. One includingthe lingual gyrus (peak at lingual gyrus, 0–99 −7) and a second clus-ter including the cerebellum and fusiform and lingual gyrus (peak atthe fusiform gyrus −36 −76 −11) (Fig. 2 – Panel C).
In summary, we found positive correlations between self-regulationperformance and cerebral metabolism within different sections of thecingulate cortex, superior frontal cortex and inferior temporal gyrus,and between decision-making performance and DLPFC/superior frontalmetabolism. Conversely, we found negative correlations between self-regulation performance and cerebral metabolism in the superior tem-poral gyrus, and between emotion recognition performance and cere-bral metabolism in the fusiform, cerebellum and lingual gyrus.
3.3.3. The relationship between voxel-based derived regions associatedwith neuropsychological performance and estimates of drug use
We only found a significant positive correlation between amount ofheroin used and metabolism in the left middle temporal gyrus(r=0.377) at pb0.01. By relaxing the alpha level to pb0.05, we foundpositive correlations between duration of cannabis (r=0.330) and co-caine used (r=0.296) and the right precentral gyrus, and negative cor-relations between the amount of cannabis used and the left middle
e (blue) and positive (yellow) correlations.
35 R
(yellow).
35 R
lue).
R
ve functions correlated with brain metabolism.
ot and cold executive functions in polysubstance addiction: Associ-Neuroimaging (2012), doi:10.1016/j.pscychresns.2012.01.006

T
359
360
361
362
363
364
365
366
367
368
369
370
371
372
373
374
375
376
377
378
379
380
381
382
383
384
385
386
387
388
389
390
391
392
393
394
395
396
397
398
399
400
401
402
403
404
405
406
407
408
409
410
411
412
413
414
415
416
417
418
419
420
421
422
423
424
425
426
427
428
429
430
431
432
433
434
435
436
437
438
439
440
441
442
443
444
445
446
447
448
449
450
451
452
453
454
455
456
457
458
459
460
461
462
463
464
465
466
467
468
469
470
471
472
473
474
475
476
477
478
479
480
481
482
483
484
485
486
487
488
6 L. Moreno-López et al. / Psychiatry Research: Neuroimaging xxx (2012) xxx–xxx
UNCO
RREC
frontal gyrus (r=−0.370) and left inferior parietal lobe (r=−0.307),the amount of heroin used and the right inferior temporal gyrus (r=−0.308), and the amount of alcohol used and the right middle frontalgyrus (r=−0.297).
4. Discussion
This study investigated the association between the neuropsycho-logical performance of SDI on measures of cold and hot executivefunctions (as measured outside the scanner) and PET-indexed region-al brain glucose metabolism. SDI were also compared to non-drugusing controls with respect to behavioral performance in these tasks.
We found that SDI had significantly poorer performance than HCindividuals on all of the neuropsychological tests administered. Like-wise, the neuropsychological performance in the SDI was just affectedby the duration of abstinence in the LNS task, allowing us to rule outthe effects of this variable on all other tasks. On the other hand, in ac-cordance with our initial hypotheses, we found that both cold and hotexecutive tests were significantly associated with common brain re-gions, including the DLPFC, mid-superior frontal gyrus and superior,inferior and middle temporal gyrus. All these regions are thought toplay important roles in the executive control of goal-driven behavior(Bechara and Van der Linden, 2005), and in neuroadaptations relatedto addiction (Koob and Volkow, 2010); therefore, our findings high-light the relevance of this set of regions for a wide range of cognitivefunctions that are significantly impaired in SDI. In terms of differentialassociations, cold executive tests mainly correlated with frontal, tem-poral and parietal regions, whereas hot executive functions correlatedwith midline mid-superior frontal and cingulate regions in both ROIand whole brain voxel-based analyses. These results are in agreementwith the notion that different executive processes rely in both com-mon and partially dissociable neural networks (Collette et al., 2006),and that different neural circuits are relevant to various aspects of ab-normal cognitive functioning in SDI (Verdejo-García et al., 2004).
Specifically, our results showed that verbal working memory(indexed by LNS) is mainly associated with DLPFC and temporo-parietal regions (Fig. 1 –Panel A), which is consistent with previousfindings suggesting these regions are the key neural substrates formemory updating and manipulation (Collette and Van der Linden,2002; D'Esposito, 2007). For response inhibition, results showed asso-ciations between Stroop performance and DLPFC and basal gangliametabolism (Fig. 1 –Panel B). This finding is in agreement with evi-dence from cognitive and neuroimaging studies showing thatfronto-striatal alterations related to inhibitory control are ubiquitousacross different forms of addiction, and are associated with difficultiesin ceasing drug use (Feil et al., 2010). For cognitive flexibility results,we showed associations between the number of errors and DLPFCand parietal cortex resting metabolism (Fig. 1 –Panel C), replicatingprevious findings using this probe and other switching-perseveration tests (e.g., the Wisconsin Card Sorting Test) in alco-holics (Adams et al., 1993) and healthy subjects (Thomas et al., 2011).
With regard to hot executive functions, results showed that R-SATperformance was associated with resting metabolism in the mid-posterior and ACC, insula and temporal regions (Fig. 2 –Panel A).Both insula and ACC are involved in task-level control and self-regulation of behavior (Posner et al., 2007; Nelson et al., 2010), thekey cognitive processes needed to develop and implement goal-driven adjustment of performance (as indexed by this task). ACCand insula dysfunctions have been associated with stress-inducedcraving, poor inhibitory control, impaired insight and less motivationto change in SDI (Sinha and Li, 2007; Li and Sinha, 2008; Goldstein etal., 2009; Romero et al., 2010); this range of symptoms may relate tocore deficits of self-regulation (i.e., difficulties to set and adjust be-havior according to long-term goals) in addicted individuals. Further-more, both anterior and posterior cingulate regions have beenpreviously associated with abnormal performance on multi-tasking
Please cite this article as: Moreno-López, L., et al., Neural correlates of hation between neuropsychological performance..., Psychiatry Research:
ED P
RO
OF
measures, specifically with deficits in task learning and remembering,rule-breaking and task switching (Burgess et al., 2000).
According to previous studies using similar methodologies, IGT per-formance in mid-term abstinent SDI is associated with resting metabo-lism in both OFC and DLPFC (Adinoff et al., 2003; Tucker et al., 2004;Krain et al., 2006) (Fig. 2 – Panel B). This finding is indicative of the no-tion that both cold processes, such as working memory or switchingskills relying on DLPFC functioning, and hot processes, such asmonitor-ing of reward value, relying on OFC functioning play a role in decision-making performance in SDI (Bolla et al., 2003). Finally, although wefailed to find significant associations between ventral fronto-striatal re-gions and emotional perception, we found robust associations betweenscores in this task and two areas (lingual and fusiform gyrus) (Fig. 2 –
Panel C) that have been consistently associated with face processingin numerous studies (Kanwisher and Yovel, 2006; Dinkelacker et al.,2010). The association between emotion processing and the ventrome-dial frontal stream could be better grasped by fMRI studies monitoringbrain activation during actual emotional arousal.
Finally, we found significant associations between patterns of druguse and the regions associated with executive performance in bothROI andwhole brain voxel-based analysis. Althoughmostly exploratory,some of these effects are informative about the link between the use ofparticular substances and the brain regions supporting different execu-tive functions. For example, higher metabolism in the middle temporalgyrus was associated with both R-SAT performance and severity of her-oin use, in agreement with data from resting fMRI showing left middletemporal gyrus dysfunction in chronic heroin users (Jiang et al., 2011).Higher metabolism in the right precentral gyrus was associated withboth severity of cocaine use and cognitive flexibility, such that we mayspeculatively suggest a role of this region in cocaine-inducedmotor per-severation deficits (Ersche et al., 2008). Lower metabolism in the supe-rior frontal gyrus was associated with both severity of alcohol use andperformance on cold executive tasks andon decision-making; this resultis fitting with the findings of Goldstein et al. (2004) who found that se-verity of alcohol use was the main predictor of a composite index of ex-ecutive impairment, whichwas predicted by regional metabolism in theanterior cingulate. Finally, lower metabolism in the adjacent middlefrontal gyrus was linked to severity of cannabis use and performanceon response inhibition and decision-making, fittingwith evidence of im-pairment of these functions in chronic cannabis users (Verdejo-Garcia etal., 2007; Battisti et al., 2010). These associations are in agreement withthe notion that amount and duration of drug use impact brain function-ing in key regions for executive control, supporting the relevance of thecorrelations between executive performance and brain metabolism inthis clinical group of SDI.
Several possible limitations of our study should be considered andaddressed by future research. In the first place, the methodology usedin this study does not allow us to monitor brain activity as a direct re-sponse to neuropsychological challenges; instead it shows deficits inresting glucose metabolism that were related to test performance.Therefore, this study cannot attribute NP function to an inability of thesubstance dependent group to activate specific regions of the brain.FMRI or PET designs that involve scanning during actual test perfor-mancemay be better suited to test the neuronal substrates of executivedysfunction in SDI. On the other hand, this design allowed us to explorethe brain correlates of highly complex neuropsychological paradigms,such as the R-SAT that need to be simplified to fit them in fMRI designs.Furthermore, PET was altogether not obtained in the control group andtherefore it is not clear whether performance differences on the NPtasks were associated with actual abnormalities (i.e. compared tohealth) in glucose metabolism at rest. Future studies should examineif there are differences in these associations between a clinical groupand a control group. A fourth limitationwas the lack of estimates of nic-otine use/dependence, considering that previous studies have observedsignificant alterations in PET-indexed cerebral metabolism associatedwith nicotine use (i.e. Rose et al., 2003; Zubieta et al., 2005) and should
ot and cold executive functions in polysubstance addiction: Associ-Neuroimaging (2012), doi:10.1016/j.pscychresns.2012.01.006

T
489
490
491
492
493
494
495
496
497
498
499
500
501
502
503
504
505
506
507
508
509
510
511
512
513
514
515516517518519520521522523524525526527528529530531532533534535536537538539540541542543544545546547548549550551552553554555556557558
559560561562563564565566567568569570571572573574575576577578579580581582583584585586587588589590591592593594595596597598599600601602603604605606607608609610611612613614615616617618619620621622623624625626627628629630631632633634635636637638639640641642643644
7L. Moreno-López et al. / Psychiatry Research: Neuroimaging xxx (2012) xxx–xxx
UNCO
RREC
be assessed in any PET study of drug use. The voxel based analysesproved to be overall more sensitive than the ROI method, this is anexpected outcome because the voxel based method is used to carryout statistical tests in smaller brain units (voxels) than the ROI analysis(whole gyri). Finally, due to its correlational design, this study cannotyield conclusions about cause–effect relationships in the area of theneuropsychology of drug addiction.
Taken together, our results show that SDI have neuropsychologi-cal deficits in both cold and hot executive functions. These deficitsare associated with brain metabolism in both common and partiallydissociable neural networks encompassing frontal, parietal, temporaland basal ganglia regions.
Acknowledgements
This study has been supported with funds from the SEJ2006-08278 project from the Spanish Ministry of Education and Science,the P07-HUM-03089 project from the Council of Science and Innova-tion. Laura Moreno López is supported by the FPU grant (AP2007-03583) from the Spanish Ministry of Science and Innovation. AntonioVerdejo-García was supported by a MCINN Jose Castillejo grant in theDepartment of Experimental Psychology, University of Cambridge.Emmanuel A. Stamatakis is supported by the Stephen Erskine Fellow-ship from Queens' College, Cambridge.
Appendix A. Supplementary data
Supplementary data to this article can be found online at doi:10.1016/j.pscychresns.2012.01.006.
References
Adams, K.M., Gilman, S., Koeppe, R.A., Kluin, K.J., Brunberg, J.A., Dede, D., Berent, S.,Kroll, P.D., 1993. Neuropsychological deficits are correlated with frontal hypometa-bolism in positron emission tomography studies of older alcoholic patients. Alco-holism, Clinical and Experimental Research 17, 205–210.
Adinoff, B., Devous, M.D., Cooper, D.B., Best, S.E., Chandler, P., Harris, T., Cervin, C.A.,Cullum, C.M., 2003. Resting regional cerebral blood flow and gambling task perfor-mance in cocaine-dependent subjects and healthy comparison subjects. The Amer-ican Journal of Psychiatry 160, 1892–1894.
Anderson, S.W., Barrash, J., Bechara, A., Tranel, D., 2006. Impairments of emotion andreal-world complex behavior following childhood- or adult-onset damage to ven-tromedial prefrontal cortex. Journal of the International Neuropsychological Socie-ty 12, 224–235.
Baler, R.D., Volkow, N.D., 2006. Drug addiction: the neurobiology of disrupted self-control. Trends in Molecular Medicine 12, 559–566.
Battisti, R.A., Roodenrys, S., Johnstone, S.J., Pesa, N., Hermens, D.F., Solowij, N., 2010.Chronic cannabis users show altered neurophysiological functioning on Strooptask conflict resolution. Psychopharmacology 212, 613–624.
Bechara, A., Van Der Linden, M., 2005. Decision-making and impulse control after fron-tal lobe injuries. Current Opinion in Neurology 18, 734–739.
Bechara, A., Damasio, A.R., Damasio, H., Anderson, S.W., 1994. Insensitivity to future con-sequences following damage to the human prefrontal cortex. Cognition 50, 7–15.
Bolla, K.I., Eldreth, D.A., London, E.D., Kiehl, K.A., Mouratidis, M., Contoreggi, C.,Matochik, J.A., Kurian, V., Cadet, J.L., Kimes, A.S., Funderburk, F.R., Ernst, M., 2003.Orbitofrontal cortex dysfunction in abstinent cocaine abusers performing adecision-making task. NeuroImage 19, 1085–1094.
Bolla, K., Ernst, M., Kiehl, K., Mouratidis, M., Eldreth, D., Contoreggi, C., Matochik, J.,Kurian, V., Cadet, J., Kimes, A., Funderburk, F., London, E., 2004. Prefrontal corticaldysfunction in abstinent cocaine abusers. The Journal of Neuropsychiatry and Clin-ical Neurosciences 16, 456–464.
Brett, M., Anton, J.L., Valabregue, R., Poline, J.B., 2002. Region of interest analysis usingan SPM toolbox. NeuroImage 16, 2.
Brooks III, J.O., Rosen, A.C., Hoblyn, J.C., Woodard, S.A., Krasnykh, O., Ketter, T.A., 2009.Resting prefrontal hypometabolism and paralimbic hypermetabolism related toverbal recall deficits in euthymic older adults with bipolar disorder. The AmericanJournal of Geriatric Psychiatry 17, 1022–1029.
Burgess, P.W., Veitch, E., de Lacy Costello, A., Shallice, T., 2000. The cognitive and neu-roanatomical correlates of multitasking. Neuropsychologia 38, 848–863.
Collette, F., Van der Linden, M., 2002. Brain imaging of the central executive componentof working memory. Neuroscience and Biobehavioral Reviews 26, 105–125.
Collette, F., Hogge, M., Salmon, E., Van der Linden, M., 2006. Exploration of the neuralsubstrates of executive functioning by functional neuroimaging. Neuroscience139, 209–221.
Dao-Castellana, M.H., Samson, Y., Legault, F., Martinot, J.L., Aubin, H.J., Crouzel, C.,Feldman, L., Barrucand, D., Rancurel, G., Féline, A., Syrota, A., 1998. Frontal dysfunc-
Please cite this article as: Moreno-López, L., et al., Neural correlates of hation between neuropsychological performance..., Psychiatry Research:
ED P
RO
OF
tion in neurologically normal chronic alcoholic subjects: metabolic and neuropsy-chological findings. Psychological Medicine 28, 1039–1048.
DeFilippis, N.A., 2002. The Category Test, Computer Version, 2nd ed. Psychological As-sessment Resources, Odessa, Florida.
Del Ser, T., García-Montalvo, J.I., Martínez Espinosa, S., Delgado Villapalos, C., Bermejo,F., 1997. The estimation of premorbid intelligence in Spanish people with the“Word Accentuation Test” and its application to the diagnosis of dementia. Brainand Cognition 33, 343–356.
Desmond, J.E., Chen, S.H., DeRosa, E., Pryor, M.R., Pfefferbaum, A., Sullivan, E.V., 2003.Increased frontocerebellar activation in alcoholics during verbal working memory:an fMRI study. NeuroImage 19, 1510–1520.
D'Esposito, M., 2007. From cognitive to neural models of working memory. Philosoph-ical Transactions of the Royal Society B: Biological Sciences 29, 761–772.
Dinkelacker, V., Grüter, M., Klaver, P., Grüter, T., Specht, K., Weis, S., Kennerknecht, I., Elger,C.E., Fernandez, G., 2010. Congenital prosopagnosia: multistage anatomical and func-tional deficits in face processing circuitry. Journal of Neurology 258, 770–782.
Eldreth, D.A., Matochik, J.A., Cadet, J.L., Bolla, K.I., 2004. Abnormal brain activity in pre-frontal brain regions in abstinent marijuana users. NeuroImage 23, 914–920.
Ersche, K.D., Roiser, J.P., Robbins, T.W., Sahakian, B.J., 2008. Chronic cocaine but notchronic amphetamine use is associated with perseverative responding in humans.Psychopharmacology 197, 421–431.
Feil, J., Sheppard, D., Fitzgerald, P.B., Yücel, M., Lubman, D.I., Bradshaw, J.L., 2010. Addic-tion, compulsive drug seeking, and the role of frontostriatal mechanisms in regu-lating inhibitory control. Neuroscience and Biobehavioral Reviews 35, 248–275.
Fernández-Serrano, M.J., Pérez-García, M., Verdejo-García, A., 2011. What are the spe-cific vs. generalized effects of drugs of abuse on neuropsychological performance?Neuroscience and Biobehavioral Reviews 35, 377–406.
Forman, S.D., Dougherty, G.G., Casey, B.J., Siegle, G.J., Braver, T.S., Barch, D.S., Stenger, V.A.,Wick-Hull, C., Pisarov, L.A., Lorensen, E., 2004. Opiate addicts lack error-dependentactivation of rostral anterior cingulate. Biological Psychiatry 55, 531–537.
Gilman, S., Adams, K., Koeppe, R.A., Berent, S., Kluin, K.J., Modell, J.G., Kroll, P., Brunberg,J.A., 1990. Cerebellar and frontal hypometabolism in alcoholic cerebellar degenera-tion studied with positron emission tomography. Annals of Neurology 28, 775–785.
Golden, C.J., 1978. Stroop Color and Word Tets. A manual for clinical and experimentaluses. Stoelting Co, Wood Dale, Illinois.
Goldstein, R.Z., Volkow, N.D., 2002. Drug addiction and its underlying neurobiologicalbasis: neuroimaging evidence for the involvement of the frontal cortex. The Amer-ican Journal of Psychiatry 159, 1642–1652.
Goldstein, R.Z., Leskowjan, A.C., Hoff, A.L., Hitzemann, R., Bashan, F., Khalsa, S., Wang,G.J., Fowler, J.S., Volkow, N.D., 2004. Severity of neuropsychological impairmentin cocaine and alcohol addiction: association with metabolism in the prefrontalcortex. Neuropsychologia 42, 1447–1458.
Goldstein, R.Z., Craig, A.D., Bechara, A., Garavan, H., Childress, A.R., Paulus, M.P.,Volkow, N.D., 2009. The neurocircuitry of impaired insight in drug addiction.Trends in Cognitive Sciences 9, 372–380.
Halpern, J.H., Pope Jr., H.G., Sherwood, A.R., Barry, S., Hudson, J.I., Yurgelun-Todd, D.,2004. Residual neuropsychological effects of illicit 3,4-methylenedioxymetham-phetamine (MDMA) in individuals with minimal exposure to other drugs. Drugand Alcohol Dependence 75, 135–147.
Hartstra, E., Oldenburg, J.F., Van Leijenhorst, L., Rombouts, S.A., Crone, E.A., 2010. Brainregions involved in the learning and application of reward rules in a two-deckgambling task. Neuropsychologia 48, 1438–1446.
Heberlein, A.S., Padon, A.A., Gillihan, S.J., Farah, M.J., Fellows, L.K., 2008. Ventromedialfrontal lobe plays a critical role in facial emotion recognition. Journal of CognitiveNeuroscience 20, 721–733.
Jager, G., Kahn, R.S., Van Den Brink, W., Van Ree, J.M., Ramsey, N.F., 2006. Long-term ef-fects of frequent cannabis use on working memory and attention: an fMRI study.Psychopharmacology 185, 358–368.
Jiang, G.H., Qiu, Y.W., Zhang, X.L., Han, L.J., Lv, X.F., Li, L.M., Lin, C.L., Zhuo, F.Z., Hu, S.Y.,Tian, J.Z., 2011. Amplitude low-frequency oscillation abnormalities in the heroinusers: a resting state fMRI study. NeuroImage 57, 149–154.
Kanwisher, N., Yovel, G., 2006. The fusiform face area: a cortical region specialized forthe perception of faces. Philosophical Transactions of the Royal Society of London.Series B, Biological Sciences 361, 2109–2128.
Kerr, A., Zelazo, P.D., 2004. Development of “hot” executive function: the children'sgambling task. Brain and Cognition 55, 148–157.
Koob, G.F., Volkow, N.D., 2010. Neurocircuitry of addiction. Neuropsychopharmacology35, 217–238.
Krain, A.L., Wilson, A.M., Arbuckle, R., Castellanos, F.X., Milham, M.P., 2006. Distinctneural mechanisms of risk and ambiguity: a meta-analysis of decision-making.NeuroImage 32, 477–484.
Lee, T.M., Zhou, W.H., Luo, X.J., Yuen, K.S., Ruan, X.Z., Weng, X.C., 2005. Neural activityassociated with cognitive regulation in heroin users: a fMRI study. NeuroscienceLetters 382, 211–216.
Levine, B., Dawson, D., Boutet, I., Schwartz, M.L., Stuss, D.T., 2000. Assessment of strate-gic self-regulation in traumatic brain injury: its relationship to injury severity andpsychosocial outcome. Neuropsychology 14, 491–500.
Li, C.S., Sinha, R., 2008. Inhibitory control and emotional stress regulation: neuroimag-ing evidence for frontal-limbic dysfunction in psycho-stimulant addiction. Neuro-science and Biobehavioral Reviews 32, 581–597.
López-Torrecillas, F., Godoy, J.F., Pérez-García, M., Godoy, D., Sánchez-Barrera, M., 2001.Variables modulating stress and coping that discriminate drug consumers fromlow or not drug consumers. Addictive Behaviors 25, 161–165.
Lubman, D.I., Yücel, M., Kettle, J.W., Scaffidi, A., Mackenzie, T., Simmons, J.G., Allen, N.B.,2009. Responsiveness to drug cues and natural rewards in opiate addiction: asso-ciations with later heroin use. Archives of General Psychiatry 66, 205–212.
ot and cold executive functions in polysubstance addiction: Associ-Neuroimaging (2012), doi:10.1016/j.pscychresns.2012.01.006

645646647648649650651652653654655656657658659660661662663664665666667668669670671672673674675676677678
679680681682683684685686687688689690691692693694695696697698699700701702703704705706707708709710711
712
713
8 L. Moreno-López et al. / Psychiatry Research: Neuroimaging xxx (2012) xxx–xxx
Lundqvist, T., 2010. Imaging cognitive deficits in drug abuse. Current Topics in Behav-ioral Neurosciences 3, 247–275.
Marinkovic, K., Oscar-Berman, M., Urban, T., O'Reilly, C.E., Howard, J.A., Sawyer, K.,Harris, G.J., 2009. Alcoholism and dampened temporal limbic activation to emo-tional faces. Alcoholism, Clinical and Experimental Research 33, 1880–1892.
Nelson, S.M., Dosenbach, N.U., Cohen, A.L., Wheeler, M.E., Schlaggar, B.L., Petersen, S.E.,2010. Role of the anterior insula in task-level control and focal attention. BrainStructure & Function 214, 669–680.
Passetti, F., Clark, L., Mehta, M.A., Joyce, E., King, M., 2008. Neuropsychological predictorsof clinical outcome in opiate addiction. Drug and Alcohol Dependence 94, 82–91.
Paulus, M.P., Tapert, S.F., Schuckit, M.A., 2005. Neural activation patterns ofmethamphetamine-dependent subjects during decision making predict relapse.Archives of General Psychiatry 62, 761–768.
Posner, M.I., Rothbart, M.K., Sheese, B.E., Tang, Y., 2007. The anterior cingulate gyrusand the mechanism of self-regulation. Cognitive, Affective, & Behavioral Neurosci-ence 7, 391–395.
Romero, M.J., Asensio, S., Palau, C., Sanchez, A., Romero, F.J., 2010. Cocaine addiction:diffusion tensor imaging study of the inferior frontal and anterior cingulatewhite matter. Psychiatry Research 181, 57–63.
Rose, J.E., Behm, F.M., Westman, E.C., Mathew, R.J., London, E.D., Hawk, T.C., Turkington,T.G., Coleman, R.E., 2003. PET studies of the influences of nicotine on neural sys-tems in cigarette smokers. The American Journal of Psychiatry 160, 323–333.
Salloum, J.B., Ramchandani, V.A., Bodurka, J., Rawlings, R., Momenan, R., George, D.,Hommer, D.W., 2007. Blunted rostral anterior cingulate response during a simpli-fied decoding task of negative emotional facial expressions in alcoholic patients.Alcoholism, Clinical and Experimental Research 31, 1490–1504.
Sinha, R., Li, C.S., 2007. Imaging stress- and cue-induced drug and alcohol craving: asso-ciation with relapse and clinical implications. Drug and Alcohol Review 26, 25–31.
Thomas, L.A., Hall, J.M., Skup, M., Jenkins, S.E., Pine, D.S., Leibenluft, E., 2011. A develop-mental neuroimaging investigation of the change paradigm. Developmental Sci-ence 14, 148–161.
Tomasi, D., Goldstein, R.Z., Telang, F., Maloney, T., Alia-Klein, N., Caparelli, E.C., Volkow,N.D., 2007. Widespread disruption in brain activation patterns to a working mem-ory task during cocaine abstinence. Brain Research 1171, 83–92.
UNCO
RRECT
Please cite this article as: Moreno-López, L., et al., Neural correlates of hation between neuropsychological performance..., Psychiatry Research:
PRO
OF
Tucker, K.A., Potenza, M.N., Beauvais, J.E., Browndyke, J.N., Gottschalk, P.C., Kosten, T.R.,2004. Perfusion abnormalities and decision making in cocaine dependence. Biolog-ical Psychiatry 56, 527–530.
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello, F., Etard, O., Delcroix, N.,Mazoyer, B., Joliot, M., 2002. Automated anatomical labelling of activations in SPMusing a macroscopic anatomical parcellation of the MNI MRI single subject brain.NeuroImage 15, 273–289.
Verdejo-García, A., Bechara, A., 2009. A somatic marker theory of addiction. Neurophar-macology 56, 48–62.
Verdejo-García, A., López-Torrecillas, F., Orozco, C., Pérez-García, M., 2004. Clinical impli-cations and methodological challenges in the study of the neuropsychological corre-lates of cannabis, stimulant and opioid abuse. Neuropsychology Review 14, 1–41.
Verdejo-Garcia, A., Benbrook, A., Funderburk, F., David, P., Cadet, J.L., Bolla, K.I., 2007.The differential relationship between cocaine use and marijuana use on decision-making performance over repeat testing with the Iowa Gambling Task. Drug andAlcohol Dependence 90, 2–11.
Verdejo-García, A., Rivas-Pérez, C., Vilar-López, R., Pérez-García, M., 2007. Strategic self-regulation, decision-making and emotion processing in poly-substance abusers intheir first year of abstinence. Drug and Alcohol Dependence 12, 139–146.
Wechsler, D., 1997. Wechsler adult intelligence scale, 3rd ed. The Psycological Corpora-tion, San Antonio, Texas.
Yaouhi, K., Bertran, F., Clochon, P., Mézenge, F., Denise, P., Foret, J., Eustache, F.,Desgranges, B., 2009. A combined neuropsychological and brain imaging study ofobstructive sleep apnea. Journal of Sleep Research 18, 36–48.
Young, A.W., Perrett, D.I., Calder, A.J., Sprengelmeyer, R., Ekman, P., 2002. Facial expressionsof emotion: stimuli and tests (FEEST). Thames Valley Test Company, Bury St. Edmunds.
Zakzanis, K.K., 2001. Statistics to tell the truth, the whole truth, and nothing but thetruth: formulae, illustrative numerical examples, and heuristic interpretation of ef-fects size analyses for neuropsychological researchers. Archives of Clinical Neuro-psychology 16, 653–677.
Zubieta, J.K., Heitzeg, M.M., Xu, Y., Koeppe, R.A., Ni, L., Guthrie, S., Domino, E.F., 2005.Regional cerebral blood flow responses to smoking in tobacco smokers after over-night abstinence. The American Journal of Psychiatry 162, 567–577.
EDot and cold executive functions in polysubstance addiction: Associ-Neuroimaging (2012), doi:10.1016/j.pscychresns.2012.01.006