actividad anti-inflamatoria in vitro de los...
TRANSCRIPT

ACTIVIDAD ANTI-INFLAMATORIA IN VITRO DE LOS EXTRACTOS Y
FRACCIONES OBTENIDAS DE LA CORTEZA INTERNA DE Tabebuia
chrysantha (JACQ.) G.NICHOLSON.
Presentado por:
STEFANYA VELÁSQUEZ GÓMEZ
VANESSA POSADA TABARES
UNIVERSIDAD TECNOLÓGICA DE PEREIRA
FACULTAD DE TECNOLOGÍA
ESCUELA DEQUÍMICA
GRUPO POLIFENOLES
Pereira, Febrero 1 de 2013

GRUPO POLIFENOLES
ACTIVIDAD ANTI-INFLAMATORIA IN VITRO DE LOS EXTRACTOS Y FRACCIONES OBTENIDAS DE LA CORTEZA INTERNA DE Tabebuia
chrysantha (JACQ.) G.NICHOLSON.
TRABAJO DE GRADO
Requisito parcial para optar al título de Químico Industrial
Presentado por:
STEFANYA VELÁSQUEZ GÓMEZ
VANESSA POSADA TABARES
Directora:
Dra. LUZ ANGELA VELOZA CASTIBLANCO
Codirector:
Dr. JUAN CARLOS SEPÚLVEDA ARIAS
UNIVERSIDAD TECNOLÓGICA DE PEREIRA
FACULTAD DE TECNOLOGÍA
ESCUELA DE QUÍMICA
GRUPO POLIFENOLES
Pereira, Febrero 1 de 2013

GRUPO POLIFENOLES
NOTA DE ACEPTACIÓN DEL TRABAJO DE GRADO
ACTIVIDAD ANTI-INFLAMATORIA IN VITRO DE LOS EXTRACTOS Y
FRACCIONES OBTENIDAS DE LA CORTEZA INTERNA DE Tabebuia
chrysantha (JACQ.) G.NICHOLSON.
Presentado por:
STEFANYA VELÁSQUEZ GÓMEZ
VANESSA POSADA TABARES
Los suscritos director y jurado del presente trabajo de grado, una vez
revisada la versión escrita y presenciado la sustentación oral, decidimos
otorgar la nota de : ______________________
Con la connotación: ______________________
Para constancia firmamos en la ciudad de Pereira hoy
_________________________________.
El director: _________________________________
Nombre: Luz Angela Veloza Castiblanco
Jurado: ________________________________
Nombre: Carlos Alberto Isaza Mejía

GRUPO POLIFENOLES
DEDICATORIA
Les dedicamos este trabajo a nuestras familias por todo el apoyo incondicional
que nos dieron para la realización de este proyecto, por su empeño, dedicación y
constancia que nos permito culminar nuestros estudios.
Stefanya Velásquez Gómez
Vanessa Posada Tabares

GRUPO POLIFENOLES
AGRADECIMIENTOS
Le agradecemos a nuestra familia por el apoyo incondicional que nos brindaron
durante toda nuestra etapa universitaria.
A la vicerrectoría de investigaciones innovación y extensión de la Universidad
Tecnológica de Pereira, por la financiación parcial de este proyecto.
A la Dra. Luz Angela Veloza Castiblanco y al Dr. Juan Carlos Sepúlveda Arias
por habernos permitido desarrollar este proyecto.
A Iván Alberto Lopera Castrillón por el apoyo constante para realizar el
presente trabajo.
A los integrantes del Grupo Polifenoles y el Laboratorio de Fisiología Celular e
Inmunología por toda la compañía y por hacer más agradable el trabajo.
A los trabajadores del jardín botánico de la UTP por su ayuda en la recolección
del material vegetal.
A la oficina de relaciones internacionales de la Universidad Tecnológica de
Pereira, por la financiación parcial del viaje a la Universidad Nacional
Autónoma de México.
Al Dr. Leovigildo Quijano por habernos dado la oportunidad de realizar la
estancia de investigación en el Instituto de Química de la UNAM y
complementar este proyecto.
A nuestros amigos por darnos momentos felices y apoyarnos durante todo este
proceso.

GRUPO POLIFENOLES
TABLA DE CONTENIDO
1 ANTECEDENTES 1
1.1 Surgimiento del problema 1
1.2 Formulación del problema 2
1.3 Objetivos 2
1.3.1 Objetivo general 2
1.3.2 Objetivos específicos 2
1.4 Justificación 3
2 MARCO TEÓRICO 7
2.1 Generalidades de la especie Tabebuia chrysantha 7
2.1.1 Usos y propiedades de la especie Tabebuia chrysantha 8
2.1.2 Compuestos químicos presentes en Tabebuia chrysantha 9
2.2 Inflamación 11
2.2.1 Tipos de inflamación 12
2.2.2 Fases de la inflamación 13
2.2.3 Mediadores de la inflamación 13
2.3 Técnicas para detectar la producción de mediadores de inflamación 18
2.3.1 Prueba de ELISA 18
2.3.2 Reacción en cadena de la polimerasa con transcripción reversa (RT-
PCR) 19
2.4 Anti-inflamatorios no esteroidales (AINES) 21
3 SECCIÓN EXPERIMENTAL 22
3.1 Materiales, equipos y reactivos 22

GRUPO POLIFENOLES
3.1.1 Material vegetal y biológico 22
3.1.2 Equipos 22
3.1.3 Reactivos 23
3.2 Obtención de los extractos 24
3.3 Fraccionamiento del extracto en cloroformo 26
3.4 Perfiles cromatográficos por HPLC 27
3.5 Marcha fitoquímica de la corteza interna de Tabebuia chrysantha 28
3.6 Ensayos biológicos 30
3.6.1 Test de viabilidad celular 30
3.6.2 Determinación de la producción de PGE2 y TNF-α 31
3.6.3 Determinación de la producción de NO 32
3.6.4 Extracción de RNA y análisis por medio de RT-PCR (transcripción
reversa reacción en cadena de la polimerasa) 32
3.6.5 Cuantificación de RNA 33
3.6.6 Electroforesis de los productos de RT-PCR 33
3.7 Fraccionamiento de la fracción F-16 del extracto en cloroformo 33
3.8 Purificación de compuestos de la F-16-3 de la fracción F-16 34
3.9 Espectroscopia de resonancia magnética nuclear (RMN) 34
3.10 Análisis estadístico 35
4 RESULTADOS Y DISCUSIÓN 36
4.1 Rendimiento de extracción 36
4.2 Perfiles cromatográficos por HPLC de los extractos de corteza interna de
T. chrysantha 37
4.3 Marcha fitoquímica 39

GRUPO POLIFENOLES
4.4 Ensayo de viabilidad celular 41
4.5 Actividad anti-inflamatoria in vitro de los extractos de T.chrysantha. 42
4.5.1 Inhibición en la producción de PGE2 42
4.5.2 Inhibición en la producción de TNF-α 43
4.5.3 Inhibición en la producción de óxido nítrico (NO) 44
4.6 CUANTIFICACIÓN DE mRNA EN LA LINEA CELULAR RAW264.7
ESTIMULADAS CON LPS 45
4.7 Efecto de los extractos sobre la expresión de mRNA de COX-2, TNFα e
iNOS en macrófagos murinos RAW264.7 estimulados con LPS. 46
4.8 Rendimiento del fraccionamiento del extracto en cloroformo. 49
4.9 Viabilidad celular de las fracciones del extracto en cloroformo 50
4.10 Efecto de las fracciones del extracto en cloroformo de T. chrysantha
sobre la producción de PGE2, TNF-α y NO 52
4.11 Rendimiento del fraccionamiento de la fracción F16 del extracto en
cloroformo 54
4.12 Purificación de compuestos de la fracción F16-3 54
5 CONCLUSIONES 58
6 RECOMENDACIONES 60
7 BIBLIOGRAFIA 61

GRUPO POLIFENOLES
INDICE DE TABLAS
Tabla 3.1. Condiciones del HPLC-DAD para fracciones polares. .......................... 27
Tabla 3.2. Pruebas de caracterización de compuestos químicos. ......................... 29
Tabla 4.1. Resultados de la marcha fitoquímica del extracto de corteza interna de
T. Chrysantha. ....................................................................................................... 40
Tabla 4.2. Cuantificación de mRNA de los macrofagos RAW264.7 estimulados con
LPS. ....................................................................................................................... 46
Tabla 4.3. Rendimiento del fraccionamiento del extracto en cloroformo ................ 50
Tabla 4.4. Viabilidad celular de los macrófagos RAW264.7 frente a las fracciones
del extracto en cloroformo de T.chrysantha. .......................................................... 51
Tabla 4.5. Rendimiento del fraccionamiento de la fracción F16. ............................ 54

GRUPO POLIFENOLES
INDICE DE FIGURAS
Figura 2.1. Compuestos químicos de la familia Bignoniaceae. ................................ 7
Figura 2.2. Tabebuia chrysantha ............................................................................. 8
Figura 2.3. Compuestos químicos de Tabebuia spp. ............................................. 10
Figura 2.4. Naftoquinonas presentes en T chrysantha. ......................................... 10
Figura 2.5. Células y matriz del tejido conjuntivo implicado en la respuesta
inflamatoria. ........................................................................................................... 12
Figura 2.6. Migración de leucocitos al tejido lesionado durante la respuesta
inflamatoria. ........................................................................................................... 13
Figura 2.7. Biosíntesis del ácido araquidónico y sus metabolitos. ......................... 15
Figura 2.8. Prostaglandina E2. ............................................................................... 16
Figura 2.9. Etapas de un ensayo de ELISA. .......................................................... 19
Figura 2.10. Reacción en cadena de la polimerasa con transcriptasa reversa. ..... 20
Figura 2.11. Estructura química de algunos anti-inflamatorios. ............................. 21
Figura 3.1. Diagrama de extracción líquido – líquido de T. chrysantha. ................ 25
Figura 3.2. Fraccionamiento del extracto en cloroformo. ....................................... 26
Figura 3.3. Gradiente de ácido fosfórico–acetonitrilo. ............................................ 28
Figura 3.4. Marcha fitoquímica de la corteza interna de T. chrysantha. ................. 30
Figura 3.5. Fraccionamiento de la fracción F-16 del extracto en cloroformo ......... 34
Figura 4.1. Rendimiento de extracción líquido-líquido del extracto en metanol. .... 36
Figura 4.2. Perfiles cromatográficos del extracto en cloroformo. (A) Cromatograma,
(B) Espectro UV/VIS del pico 4 y (C) Espectro UV/VIS del pico 7. ........................ 37

GRUPO POLIFENOLES
Figura 4.3. Perfiles cromatográficos del extracto en acetato de etilo. (A)
Cromatograma, (B) Espectro UV/VIS del pico 2 y (C) Espectro UV/VIS del pico 7.
............................................................................................................................... 38
Figura 4.4. Espectro UV/VIS de la 1,4-benzoquinona. ........................................... 39
Figura 4.5. Viabilidad celular de los macrófagos murinos RAW264.7 frente a los
extractos de corteza interna de T. chrysantha. ...................................................... 41
Figura 4.6. Efecto inhibitorio de los extractos de la corteza interna de T. chrysantha
sobre la producción de PGE2 en macrófagos murinos estimulados con LPS. ....... 43
Figura 4.7. Efecto inhibitorio de los extractos de la corteza interna de T. chrysantha
sobre la producción de TNF-α en macrófagos murinos estimulados con LPS. ...... 44
Figura 4.8. Efecto inhibitorio de los extractos de la corteza interna de T. chrysantha
sobre la producción de NO en macrófagos murinos estimulados con LPS. .......... 45
Figura 4.9. Efecto inhibitorio de las expresiones de mRNA de COX-2, TNF-α e
iNOS sobre macrófagos murinos RAW264.7 estimulados con LPS. ..................... 47
Figura 4.10. Efecto inhibitorio de las fracciones del extracto en cloroformo de la
corteza interna de T. chrysantha sobre la producción de PGE2 (A), TNF-α (B) y NO
(C) en macrófagos murinos estimulados con LPS. ................................................ 53
Figura 4.11. Espectro de RMN de 1H de la fracción F16-3-b ................................. 55
Figura 4.12. Espectro de RMN de 13C de la fracción F16-3-b ................................ 56
Figura 4.13.Posible estructura de compuesto mayoritario de la fracción F16-3-b . 57
Figura 4.14. Espectro de RMN de 1H del ácido veratrico. ...................................... 57

GRUPO POLIFENOLES
RESUMEN
Tabebuia chrysantha es una especie perteneciente a la familia Bignoniaceae, es
un árbol de hasta 35 m de altura, la corteza es de color gris a café oscuro, y las
flores son grandes de color amarillo. Esta especie está distribuida en
Centroamérica y en parte de Sudamérica. Es apreciada por su belleza ornamental,
por la dureza de su madera y se reportan diversos usos en medicina tradicional
como antifúngico, antitumoral y anti-inflamatorio. Estas actividades son similares a
las encontradas en otras especies pertenecientes a la familia Bignoniaceae como
Tabebuia impetiginosa, Tabebuia rosea, Tabebuia aurea, entre otras.
En el presente estudio se evaluó la actividad anti-inflamatoria in vitro de los
extractos en n-hexano, cloroformo, acetato de etilo, butanol y agua obtenidos a
partir de la corteza interna de Tabebuia chrysantha. La marcha fitoquímica
realizada a partir del material vegetal muestra la presencia de glicósidos
cardiotónicos, quinonas y lactonas sesquiterpenicas.
Se realizaron ensayos de viabilidad celular para evaluar los efectos citotóxicos de
los extractos a tres concentraciones (2,5 µg/mL, 1,2 µg/mL y 0,6 µg/mL). La
concentración de 2,5 µg/mL mostró porcentajes de viabilidad celular superiores al
80% y por lo tanto se empleó esta concentración para estimular macrófagos
murinos (RAW264.7) con Lipopolisacárido bacteriano (LPS), en presencia de los
extractos vegetales y se determinó la producción de Prostaglandina E2 (PGE2),
Factor de Necrosis Tumoral alfa (TNF-α) y Óxido Nítrico (NO).
El extracto en cloroformo mostró efecto anti-inflamatorio in vitro, inhibiendo la
producción de PGE2 y TNF-α, constituyéndose en el extracto más promisorio de
todos los evaluados. Dicho extracto fue fraccionado mediante cromatografía en
columna, obteniéndose 21 fracciones a las cuales se les evaluó su efecto sobre la
viabilidad celular de células RAW264.7. La evaluación de la actividad anti-
inflamatoria de las fracciones con mayor rendimiento (F12, F13 y F16) mostró que
las fracciones F13 y F16 inhibieron la producción tanto de PGE2 como de NO.

GRUPO POLIFENOLES
La fracción F16 fue fraccionada mediante cromatografía en columna,
obteniéndose 9 fracciones de las cuales se decidió purificar la fracción F16-3 por
cromatografía preparativa, las fracciones resultantes fueron analizadas por
espectroscopia de resonancia magnética nuclear (RMN) de 1H y 13C.

GRUPO POLIFENOLES
ABSTRACT
Tabebuia chrysantha belongs to the family Bignoniaceae, is a tree of 35 m high,
the bark is gray to dark brown, and the flowers are bright yellow. This species are
distributed in Central America and parts of South America. It is treasure for its
ornamental beauty, because of the hardness of their wood and reported several
uses in traditional medicine as antifungal, antitumor and anti-inflammatory. These
activities are similar to those found in other species that belong to the family
Bignoniaceae as Tabebuia impetiginosa, Tabebuia rosea, Tabebuia aurea.
In the present study we assessed the anti-inflammatory activity in vitro of the
extracts in n-hexane, chloroform, ethyl acetate, butanol and water derived from the
inner bark of Tabebuia chrysantha. The phytochemical study of the plant material
shows the presence of cardiac glycosides, quinones and sesquiterpene lactones
Cell viability assays were performed to test cytotoxic effects of the extracts at three
concentrations (2.5 µg / mL, 1.2 µg / mL and 0.6 µg / mL). The concentration of 2.5
µg / mL exhibits cell viability over 80% this concentration was used to stimulate
murine macrophages (RAW264.7) with bacterial lipopolysaccharide (LPS) in the
presence of plant extracts and determined the production of prostaglandin E2
(PGE2), tumor necrosis factor alpha (TNF-α) and nitric oxide (NO).
The chloroform extract showed anti-inflammatory effects in vitro, inhibiting the
production of PGE2 and TNF-α, and it becomes the most promising extract
evaluated. This extract was fractionated by chromatography column to give 21 Cell
viability assays were performed to test cytotoxic effects of the fractions F13 and
F16 inhibited PGE2 production and NO.
Fraction F16 was fractionated by chromatography column and getting 9 fractions.
F16-3 was purified by chromatography preparative; the resulting fractions were
analyzed by nuclear magnetic resonance spectroscopy (NMR) 1H and 13C.

GRUPO POLIFENOLES
INTRODUCCIÓN
A lo largo de los años las plantas han sido usadas para el tratamiento de diversas
enfermedades, incluyendo aquellas de origen inflamatorio y es por esta razón que
en el presente trabajo se pretende brindar información sobre la actividad anti-
inflamatoria de Tabebuia chrysantha.
Las especies pertenecientes al género Tabebuia tienen usos etnomédicos en
Sudamérica, en especial la especie Tabebuia impetiginosa, a la cual se la ha
demostrado actividades citotóxica y antitumoral, inmunosupresora, anti-
inflamatoria, inmunomoduladora, anti-ulceroso, antibacterial y antioxidante, entre
otras, convirtiéndose así en un referente para la presente investigación.
En el caso específico de T. chrysantha el extracto de su corteza interna es
considerado como agente antirreumático, se utiliza para el tratamiento de úlceras,
infecciones crónicas y alergias. Se ha demostrado que esta especie presenta
actividad antifúngica, larvicida y anti-inflamatoria. A partir de esta especie se han
aislado compuestos como antraquinonas, naftoquinonas y furanoquinonas.
Las enfermedades neurodegenerativas e inflamatorias son actualmente un
problema de salud pública a nivel mundial. Debido al hecho de que dichas
enfermedades poseen algunos mecanismos fisiopatológicos similares, se ha
decidido buscar alternativas terapéuticas en plantas de la familia Bignoniaceae,
específicamente en Tabebuia chrysantha, la cual ha sido poco estudiada tanto
desde el punto de vista de sus constituyentes como de sus actividades biológicas
y en especial, la actividad anti-inflamatoria. Se pretende obtener compuestos que
tengan actividad anti-inflamatoria, que puedan ser promisorios como nuevos
medicamentos. La identificación de moléculas promisorias como anti-inflamatorios
permitirá avanzar en la búsqueda de opciones terapéuticas para los pacientes con
enfermedades como aterosclerosis, artritis, entre otras.

GRUPO POLIFENOLES
1
1 ANTECEDENTES
1.1 Surgimiento del problema
En la actualidad los medicamentos anti-inflamatorios no esteroidales (AINEs)
causan efectos secundarios como sangrado intestinal, entre otras patologías
denominadas gastropatías (1), convirtiéndose en un problema para la salud.
En el proceso inflamatorio se han involucrado diferentes mediadores químicos que
pueden ser sintetizados por la vía de la ciclooxigenasa, de la que existen dos
isoformas denominadas COX-1 y COX-2. La primera se expresa en forma
constitutiva en casi todas las células y la COX-2 necesita ser inducida por
estímulos inflamatorios, induciendo la producción de mediadores de la inflamación
como la PGE2 (2).
Los denominados anti-inflamatorios no esteroideos (AINEs) que son utilizados
actualmente, inhiben las actividades de COX-1, COX-2 o ambas enzimas (3). La
inhibición de COX-1 lleva al desarrollo de efectos indeseables en el organismo
como las gastropatías. Los inhibidores selectivos de COX-2 disminuyen la
inflamación y son menos gastrolesivos, sin embargo el uso prolongado de estos
puede generar efectos secundarios indeseables como problemas cardiovasculares
(4).
Las enfermedades de origen inflamatorio como la artritis, el cáncer, la
aterosclerosis y la obesidad, entre otras, se consideran actualmente como
problemas importantes de salud pública y tienen como origen el desarrollo de un
proceso inflamatorio crónico y por tanto, la búsqueda de nuevas moléculas anti-
inflamatorias permitirá desarrollar nuevos medicamentos que puedan ser usados
para el tratamiento de dichas enfermedades.
A lo largo de los años las plantas han sido un recurso para el desarrollo de nuevos
medicamentos, por lo tanto, con base en hallazgos realizados en plantas del
genero Tabebuia, se pretende evaluar la actividad anti-inflamatoria de los

GRUPO POLIFENOLES
2
extractos obtenidos de la corteza interna de Tabebuia chrysantha, como una
alternativa para el desarrollo de nuevos medicamentos para el tratamiento de
procesos inflamatorios agudos y/o crónicos (2).
Algunas especies pertenecientes al género Tabebuia han demostrado tener
actividad biológica como lo son Tabebuia impetiginosa que tiene actividad
antitumoral y anti-inflamatoria (5), Tabebuia rosea que posee actividad
antimicrobiana (6), y Tabebuia chrysantha la cual se ha empleado en medicina
tradicional en Sudamérica como anti-inflamatorio, antifúngico y larvicida (7). Sin
embargo, se requiere de confirmación experimental de los usos etnomédicos de la
especie T. chrysantha, en especial, la evaluación de su efecto anti-inflamatorio.
1.2 Formulación del problema
¿Poseen los extractos y fracciones de Tabebuia chrysantha actividad inhibitoria
sobre las enzimas COX-2, iNOS y TNF-α?
1.3 Objetivos
1.3.1 Objetivo general
Determinar la actividad anti-inflamatoria in-vitro de los extractos y fracciones
obtenidas de la corteza interna de Tabebuia chrysantha.
1.3.2 Objetivos específicos
Obtener los extractos y fracciones de la corteza interna de Tabebuia
chrysantha.
Caracterizar los extractos y fracciones mediante HPLC-UV-DAD
(Cromatografía liquida de alta eficiencia con detector UV de arreglo de diodos).
Evaluar la actividad inhibitoria de COX-2, iNOS y TNF-α en los extractos y
fracciones de Tabebuia chrysantha.

GRUPO POLIFENOLES
3
1.4 Justificación
En la familia Bignoniaceae se encuentra el género Tabebuia, dentro del cual se
conocen varias especies que han sido introducidas en Colombia. En la región del
eje cafetero se encuentran las especies Tabebuia rosea (bertol.) DC. y Tabebuia
chrysantha (jacq.) Nicholson, las cuales poseen inflorescencia rosada y amarilla
respectivamente.
El género Tabebuia pertenece a áreas tropicales y subtropicales, comprende
varias especies productoras de madera, la cual es utilizada en la construcción
naval y de edificios, debido a que su madera es muy pesada, durable y resistente
(8); este género se caracteriza por sus flores de colores vistosos y el uso de su
corteza como tratamiento en medicina tradicional.
Los usos en medicina tradicional de plantas pertenecientes al género Tabebuia se
respaldan en estudios realizados en otras especies de este mismo género, como
Tabebuia impetiginosa conocida también como “lapacho”, originaria del Brasil y
norte de Argentina, la cual ha mostrado tener actividad inmunosupresora (9), anti-
inflamatoria (10), inmunomoduladora (11), anti-ulceroso (12), antibacterial (13) y
antioxidante (14). De esta especie se han aislado compuestos como quinonas,
furanonaftoquinonas, lapachol y naftoquinonas. Se ha demostrado que dichos
compuestos tienen actividad antitumoral y anti-inflamatoria (15). En el caso de
Tabebuia rosea, se ha encontrado actividad antimicrobiana contra Bacillus subtilis,
Salmonella typhimurium y Candida albicans.(7)
Tabebuia chrysantha es un árbol de hasta 35 m de altura y diámetro de hasta 60
cm, ramas escasas, gruesas y ascendentes. La corteza es áspera de color gris a
café oscuro, tiene grietas verticales, profundas y forman placas anchas de color
café oscuro. Las flores son grandes de 5 a 12 cm de largo, de color amarillo claro,
muy vistosas con líneas rojas en el cuello (16).
Existen pocos estudios fitoquímicos de la especie Tabebuia chrysantha, sin
embargo, se han aislado metabolitos secundarios de su corteza interna tales como

GRUPO POLIFENOLES
4
naftoquinonas y lapachos; además se ha demostrado que dichos compuestos
poseen actividad antifúngica (17) y larvicida (18).
Varias especies de Tabebuia tienen usos etnomédicos en Sudamérica, tal es el
caso de Tabebuia chrysantha, debido a que el extracto de su corteza interna es
considerado como agente antirreumático, se utiliza para el tratamiento de úlceras,
infecciones crónicas y alergias (19). Estas enfermedades tienen en común el
desarrollo de un proceso inflamatorio.
La inflamación básicamente se define como un cambio del equilibrio morfológico
en un área específica de un tejido, causada por diferentes agentes: físicos,
químicos, o biológicos (20). En este contexto, la respuesta inflamatoria es una
reacción crítica protectora a la irritación, lesión, o infección, caracterizada por
enrojecimiento, calor, hinchazón, pérdida de la función y dolor, a pesar de esto, los
procesos inflamatorios pueden ser perjudiciales, como ocurre en las reacciones de
hipersensibilidad luego de picaduras de insectos o en procesos inflamatorios
crónicos como la artritis reumatoidea (21).
Existe una amplia variedad de mediadores químicos implicados en el proceso
inflamatorio, dentro de los cuales se pueden incluir las aminas vasoactivas,
eicosanoides, citocinas y óxido nítrico.
Cuando las células son activadas por diversos estímulos, los lípidos de la
membrana celular generan mediadores lipídicos que van a actuar a nivel del
proceso inflamatorio. Dentro de estos lípidos, el ácido araquidónico es importante
porque da origen a los denominados eicosanoides (metabolitos del ácido
araquidónico), sintetizados mediante dos vías: la vía de la ciclooxigenasa
(generadora de prostaglandinas y tromboxanos) y la vía de la lipoxigenasas
(generadora de leucotrienos) (22).
La vía de la ciclooxigenasa es mediada por dos enzimas diferentes (COX-1 y
COX-2), denominadas isoenzimas. La COX-1 se expresa constitutivamente en
varias células, mientras que la COX-2 es inducida por estímulos proinflamatorios

GRUPO POLIFENOLES
5
como citoquinas y lipopolisacáridos, siendo muy importante en los procesos
inflamatorios (23).
La acción de la COX sobre el ácido araquidónico genera la prostaglandina H2
(PGH2), muy inestable, y precursora de otras prostaglandinas, como la PGE2 o la
PGF2α. Las prostaglandinas estimulan la inflamación, regulan el flujo sanguíneo
local y participan en la patogenia del dolor y la fiebre.(24).
El óxido nítrico (NO) es otro mediador del proceso de inflamación generado por la
isoforma inducible de la óxido nítrico sintasa (iNOS). En las células de mamíferos
se han identificado tres isoformas de esta enzima; a nivel del endotelio (eNOS), a
nivel neuronal (nNOS) y la inducida a través de un estímulo bacterial (iNOS) (25).
El NO se encarga de la vasodilatación y reduce la agregación plaquetaria, produce
a la vez radicales libres tóxicos a nivel celular (22).
Las citocinas son un grupo de proteínas producidas por distintos tipos de células
del sistema inmune en respuesta a una activación celular, entre estas se
encuentra el factor de necrosis tumoral alfa (TNF-α) (26) ,el cual se origina en los
macrófagos y actúa sobre las células endoteliales, provocando exudación de
líquidos y factores de adherencia (27).
Actualmente, las drogas anti-inflamatorias no esteroidales (AINEs) son
ampliamente empleadas en el tratamiento del dolor, fiebre y como alternativas
terapéuticas para el tratamiento de inflamaciones crónicas. Dentro de este grupo
de medicamentos, hay inhibidores de COX-1 y COX-2 en las dosis empleadas
como anti-inflamatorios. Tanto el efecto anti-inflamatorio, analgésico y antipirético
se basan en la inhibición de COX-2 pero su toxicidad gastrointestinal (asociada
con sangrado) es el resultado de la inhibición de COX-1 (28). Los efectos
desfavorables de los AINEs han restringido su uso a largo plazo, existiendo
entonces una demanda constante de nuevas opciones terapéuticas. (29)
Las enfermedades de origen inflamatorio como la artritis, el cáncer, la
aterosclerosis y la obesidad, entre otras, se consideran actualmente como

GRUPO POLIFENOLES
6
problemas importantes de salud pública y tienen como origen el desarrollo de un
proceso inflamatorio crónico y por tanto, la búsqueda de nuevas moléculas anti-
inflamatorias permitirá desarrollar nuevos medicamentos que puedan ser usados
para el tratamiento de dichas enfermedades.
Los productos naturales, incluidos los derivados de las plantas superiores, a lo
largo de los años han contribuido en gran medida al desarrollo de medicamentos.
Algunos de los metabolitos secundarios que poseen actividad anti-inflamatoria
son: terpenos, compuestos fenólicos, cumarinas y compuestos lactónicos (30),
estos metabolitos tienen efectos sobre diversos mediadores de la inflamación,
entre los cuales se encuentran los eicosanoides, generados por la acción de la
ciclooxigenasa sobre el ácido araquidónico para generar leucotrienos y
prostaglandinas (PGE2 y PGF2a) (21)
Debido a la poca información disponible con relación a los componentes y al
efecto anti-inflamatorio de los extractos vegetales de la especie T. chrysantha y
teniendo en cuenta la necesidad de identificar nuevos agentes anti-inflamatorios,
es importante iniciar estudios tendientes a evaluar la actividad biológica de dichos
extractos con el fin de determinar compuestos de origen vegetal que puedan
convertirse en una alternativa para el desarrollo de nuevos medicamentos dirigidos
al tratamiento de procesos inflamatorios agudos y/o crónicos.

GRUPO POLIFENOLES
7
2 MARCO TEÓRICO
2.1 Generalidades de la especie Tabebuia chrysantha
T. chrysantha pertenece a la familia Bignoniaceae que comprende más de 120
géneros y 650 especies distribuidas en los bosques montanos, secos y de tierras
bajas (31), primordialmente de área tropical con escasa presencia en regiones
templadas de climas cálidos.
En esta familia existen reportes de compuestos químicos (Fig. 2.1) tales como
saponinas, quinonas y taninos(32).
O
O
OR
Saponina
O
O
OH
HO
OH
OH
O OH Quinona
Tanino
Figura 2.1. Compuestos químicos de la familia Bignoniaceae.
En Colombia la familia Bignoniaceae está representada por 45 géneros nativos y
algunos introducidos por su carácter ornamental, siendo los géneros Arrabidaea y
Tabebuia los más representativos (33); este último ha sido uno de los más
estudiados, por ser una fuente de compuestos naftoquinónicos sustituidos (34) con
propiedades anticancerígenas (35), (36). Algunas especies pertenecientes al

GRUPO POLIFENOLES
8
género Tabebuia han demostrado tener actividad anti-inflamatoria (37),
antioxidante (14), antiofídica (6), antibacterial (38), entre otras.
Tabebuia chrysantha (Fig. 2.2) es un árbol conocido en Colombia como Guayacán
amarillo, el cual se distribuye geográficamente desde México hasta Venezuela. Se
desarrolla en suelos franco arenosos con alta cantidad de materia orgánica y un
pH de 6 a 8,5.
Es un árbol de hasta 35 m de altura, corteza de color gris pálido a oscuro. Las
hojas son alternas, digitadamente compuestas, de 5 a 25 cm de largo y de 8 a 20
cm de ancho, caducifolias con el haz verde oscuro y el envés verde claro y
densamente cubierto por pelos estrellados color café. Las flores son
campanuladas, grandes, de 5 a 12 cm de largo, de color amarillo claro, muy
vistosas con líneas rojas en el cuello (16).
Figura 2.2. Tabebuia chrysantha
2.1.1 Usos y propiedades de la especie Tabebuia chrysantha
El principal uso de esta especie no maderable es medicinal. Se considera que la
corteza es eficaz para tratar reumatismo, artritis, cáncer, infecciones,
inflamaciones y úlceras. Es muy apreciada como ornamental en Tarapacá

GRUPO POLIFENOLES
9
(Amazonas, Colombia) y presenta varios usos medicinales. Para curar el “meo de
arco” (sarpullido que se presenta en los niños pequeños) se raspa la corteza y el
polvillo se aplica sobre la parte afectada (39).
Estudios etnofarmacológicos realizados en personas del área rural, urbana y en
herbolarios de diferentes municipios del eje cafetero han permitido identificar las
plantas más utilizadas como antimicrobianos o estimulantes del sistema inmune,
dentro de dichas plantas se encuentra T. chrysantha (40), (41). Investigaciones
preliminares han indicado que los extractos obtenidos de las hojas de T.
chrysantha pueden tener efecto sobre la respuesta inmune al incrementar in vivo
en ratas Wistar los niveles de anticuerpos dirigidos contra glóbulos rojos de
carnero (42). Un estudio reciente reporta que tanto el extracto acuoso como el
metanólico obtenidos de las hojas de T. chrysantha poseen actividad anti-
inflamatoria in vivo (43).
2.1.2 Compuestos químicos presentes en Tabebuia chrysantha
Los principales constituyentes de los extractos de la corteza de Tabebuia spp.
incluyen furanonaftoquinonas (44), (45), (46), quinonas (47),naftoquinonas (48)
(Fig. 2.3), ácido benzóico y derivados del benzaldehido (49), ciclopenteno
dialdehidos (37), flavonoides (50) y glicósidos, principalmente del tipo iridoides
(51). Dentro de las naftoquinonas, el lapacol y la beta lapacona se han estudiado
intensivamente dadas sus propiedades antiinflamatorias (52) y anticancerígenas
(36), (53), (54), (55), (56).

GRUPO POLIFENOLES
10
O
O
O
R
O
O
R
RC
O OH
Naftoquinona
Furanonaftoquinona
Ácido bezóico
Figura 2.3. Compuestos químicos de Tabebuia spp.
Los estudios fitoquímicos llevados a cabo con la especie T. chrysantha indican
que los componentes presentes en la corteza interna son lapacol, α y β-lapacona,
dehidrotectol, tetrahidrotectol (57), entre otras naftoquinonas (58), (34) (Fig. 2.4).
O
O
OH
Lapachol
O
O
O
-lapachona
O
O
O
-lapachona
O
O
O
O
Dehidrotectol
Figura 2.4. Naftoquinonas presentes en T chrysantha.

GRUPO POLIFENOLES
11
2.2 Inflamación
La inflamación es una respuesta esencial en la supervivencia de los organismos
multiceluares frente a la agresión y consiste básicamente en una respuesta de los
vasos sanguíneos y de los leucocitos. La lesión tisular que desencadena esta
respuesta puede ser causada por un agente físico, químico o biológico. La
respuesta inflamatoria es fundamental para inducir una respuesta inmune
específica que sea adecuada y finalmente para iniciar el proceso de reparación de
los tejidos lesionados. La reparación inicia durante las fases iniciales de la
inflamación, aunque no finaliza hasta que se ha neutralizado el estímulo lesivo.
Durante la reparación, el tejido lesionado es substituido por nuevo tejido
(cicatrización).
A pesar de que el proceso inflamatorio es un mecanismo de defensa ante la
agresión tisular, muchas enfermedades como las reacciones de hipersensibilidad
o enfermedades crónicas como la artritis reumatoidea, la aterosclerosis, la fibrosis
pulmonar y la obesidad, la respuesta inflamatoria persistente induce patología y
daño tisular.
La respuesta inflamatoria tiene lugar en el tejido conectivo vascularizado (Fig. 2.5)
e involucra diferentes tipos de células como monocitos, neutrófilos, eosinófilos,
linfocitos, plaquetas, mastocitos, macrófagos y fibroblastos.

GRUPO POLIFENOLES
12
Mastocito Fibroblasto Macrófago
Fibras elásticas
Fibras de colágeno Proteoglucanos
Leucocito polimorfonuclear
Linfocito
Plaquetas
Monocito Eosinófilo
Basófilo
Células deltejido conjuntivo
Vasos
Matriz deltejido conjuntivo
Figura 2.5. Células y matriz del tejido conjuntivo implicado en la respuesta
inflamatoria.
2.2.1 Tipos de inflamación
La respuesta inflamatoria presenta dos tipos, que se diferencian por su duración
2.2.1.1 Inflamación aguda
Es la respuesta inicial e inmediata a la lesión, relacionada principalmente con
cambios hemodinámicos locales que llevan a manifestaciones clínicas como rubor,
calor, edema y dolor en la zona afectada. Posteriormente, hay salida de leucocitos
hacia el tejido lesionado e iniciar el proceso de degradación de los tejidos
necróticos. Este tipo de inflamación tiene una duración que oscila entre, minutos,
horas y pocos días.
2.2.1.2 Inflamación crónica
Es una inflamación de larga duración (semanas, meses o años) en la cual la
inflamación activa, la lesión tisular y el proceso de reparación suceden al mismo
tiempo. La inflamación crónica es nociva para la salud y se constituye en el

GRUPO POLIFENOLES
13
mecanismo patogénico básico de algunas enfermedades crónicas como artritis
reumatoidea, aterosclerosis fibrosis pulmonar, obesidad, entre otras.
2.2.2 Fases de la inflamación
Después de un corto periodo de vasoconstricción a nivel arterial, se produce
vasodilatación y aumento de flujo sanguíneo en la zona de la lesión, que causa
enrojecimiento y aumento de la temperatura; posteriormente el flujo sanguíneo
disminuye por un aumento de la permeabilidad vascular que permite la salida de
líquido de los vasos y aumento de la viscosidad sanguínea, lo que se denomina
estasis (parálisis total del flujo). A medida que evoluciona la estasis se produce la
migración de los leucocitos a través del endotelio vascular mediante un
mecanismo denominado transmigración (Fig.2.6), el cual permite que las células
inflamatorias lleguen al sitio de la lesión. (22).
Rodamiento Activación Adhesión Transmigración
Estímulo
Figura 2.6. Migración de leucocitos al tejido lesionado durante la respuesta
inflamatoria.
2.2.3 Mediadores de la inflamación
Los mediadores químicos producidos por células del sistema inmune son los
responsables del desarrollo de la respuesta inflamatoria (59). Los mediadores

GRUPO POLIFENOLES
14
inflamatorios pueden ser secretados por células especializadas del sistema
inmune y muchos de ellos son liberados a partir de gránulos intracelulares que
liberan dichos mediadores en respuesta al estímulo inflamatorio.
Como respuesta al fenómeno inflamatorio, se liberan una serie de mediadores que
funcionan de manera coordinada. Dichos mediadores se generan a partir de
células o derivan de proteínas del plasma sanguíneo. Entre los mediadores de
origen celular se tienen la histamina, serotonina, prostaglandinas, leucotrienos,
NO, citocinas (IL-1, TNF-α) y quimiocinas, entre otros. Los mediadores derivados
de proteínas plasmáticas incluyen las proteínas del complemento (C3a, C4a, C5a),
cininas y proteasas activadas durante la coagulación (22).
Para efectos de esta tesis, se mencionarán aspectos relacionados con los
mediadores evaluados en el trabajo experimental y que corresponden a
prostaglandinas, NO y citocinas.
2.2.3.1 Metabolitos del ácido araquidónico: (AA)
El ácido araquidónico es un ácido graso poliinsaturado de 20 átomos de carbono
que procede de la dieta a partir del ácido linoléico, éste se encuentra esterificado
en los fosfolípidos de la membrana celular y se libera por medio de fosfolipasas
celulares que se activan mediante estímulos proinflamatorios.
Los metabolitos del AA, llamados eicosanoides, son sintetizados mediante dos
vías: la vía de la ciclooxigenasa (generadora de prostaglandinas y tromboxanos) y
la vía de la lipooxigenasa (generadora de leucotrienos y lipoxinas) (Fig. 2.7). Las
prostaglandinas y los tromboxanos son responsables de la vasodilatación, la
generación del dolor y la fiebre. También está la vía de la lipoxigenasa que genera
leucotrienos (los cuales participan en el incremento de la permeabilidad vascular,
quimiotaxis, activación y adhesión de leucocitos) y las lipoxinas, que inhiben la
adhesión de neutrófilos y la quimiotaxis (22).

GRUPO POLIFENOLES
15
Fosfolípidos
Fosfolipasa A
Ácido araquidónico
5-Lipoxigenasa Cicloxigenasa
PGG2LTA4
LTC4
LTD4
LTD4
PGH2
PGH2
PGI2
Prostaciclina sintetasa Tromboxano sintetasa
TXA2
PGD2 PGE2 PGF2
Figura 2.7. Biosíntesis del ácido araquidónico y sus metabolitos.
La prostaglandina E2 (PGE2) (Fig. 2.8) induce vasodilatación y actúa
sinergísticamente con otros mediadores como la histamina y las bradiquininas
para incrementar la permeabilidad vascular durante la respuesta inflamatoria y
generar edema. Adicionalmente, la PGE2 es un mediador central de la respuesta
febril inducida durante el proceso inflamatorio.

GRUPO POLIFENOLES
16
O
HO
COOH
OH
Prostaglandina E2
Figura 2.8. Prostaglandina E2.
2.2.3.2 Óxido nítrico: (NO)
El oxido nítrico es un gas soluble sintetizado principalmente por vía enzimática
catalizada por la óxido nítrico sintasa (NOS) a través de una serie de reacciones
redox, con degradación de L-arginina a L-citrulina en presencia de oxígeno y
NADPH. Se conocen tres isoformas de la NOS: endotelial (eNOS o NOS3),
neuronal (nNOS o NOS1) e inducible (iNOS o NOS2). La iNOS es una enzima
inducible a nivel transcripcional y se expresa en macrófagos y otros tejidos en
respuesta a estímulos inflamatorios (60). La reacción se muestra a continuación:
NADPH, O2
L-Arg L-citrulina + NO
Tiene como función relajar y vasodilatar el musculo liso de la pared vascular y
reducir la agregación y adhesión plaquetaria (22). Adicionalmente, el NO generado
por los macrófagos tiene como función destruir microorganismos. El NO reacciona
con radicales libres del oxígeno (O2-) generando peroxinitrito, el cual tiene efectos
tóxicos sobre las células y los tejidos (61).
NO + O2 ONOO-

GRUPO POLIFENOLES
17
El peroxinitrito es un anión altamente oxidativo, capaz de oxidar tioles y las bases
del DNA, además de iniciar la peroxidación de lípidos (62).
2.2.3.3 Citocinas
Son proteínas producidas principalmente por el sistema inmune en respuesta a
una activación celular, tienen como función, regular la respuesta inmune y la
respuesta inflamatoria (26).
Las citocinas se dividen en cinco grupos dependiendo de su función: citocinas que
regulan la función leucocitaria, implicadas en la inmunidad natural, las que
estimulan la hematopoyesis, las quimiocinas y las que participan de la respuesta
inflamatoria. Las citocinas proinflamatorias por excelencia corresponden al TNF-α
y a la Interleucina-1b (IL-1β) y son producidas principalmente por los macrófagos
activados por endotoxinas como el LPS u otros productos microbianos, complejos
inmunes o daño tisular.
La principal función del TNF-α es el reclutamiento de monocitos y neutrófilos en el
sitio de entrada del agente extraño, sin embargo, hay otra gran variedad de
funciones, entre las que tenemos: estimular la capacidad de macrófagos y células
endoteliales para secretar quimiocinas y otras citocinas como la IL-1, aumentar la
capacidad microbicida de los neutrófilos, estimular la angiogénesis, coestimulación
en la activación de la célula T, inducción de fiebre (pirógeno endógeno),
incrementar la síntesis de proteínas de fase aguda como la proteína C reactiva,
inducción de muerte celular por apoptosis, inducción de caquexia, estimular la
producción del factor tisular por el endotelio activando la coagulación (22).
Con relación a la IL-1β, algunas de sus acciones son indirectas debido a la
inducción de la síntesis de otras moléculas como la PGE2, la IL-6 y la IL-8. Una de
sus principales funciones es la activación de los linfocitos T ayudadores, actúa
sinérgicamente con el TNF-α para incrementar la síntesis de proteínas de fase
aguda y activar el sistema de la coagulación. A diferencia del TNF-α, la IL-1β no
induce daño celular por apoptosis pero se induce en respuesta a LPS y potencia
los daños tisulares inducidos por el TNF- (22).

GRUPO POLIFENOLES
18
2.3 Técnicas para detectar la producción de mediadores de inflamación
A continuación se describen algunas de las técnicas empleados para la medición
de los mediadores de la inflamación como prostaglandinas, NO y citocinas.
2.3.1 Prueba de ELISA
La prueba de ELISA (Ensayo por inmunoabsorción ligado a enzimas), es una
técnica de laboratorio muy común que es utilizada para determinar una sustancia,
generalmente una antígeno, contenido en una muestra liquida. Este tipo de prueba
es comúnmente usada en medicina como herramienta para la detección de
anticuerpos y el diagnóstico de enfermedades infecciosas, sin embargo, sus usos
a nivel clínico son muy amplios. Esta técnica también se usa en la industria para
determinar la calidad de un producto.
El principio de la prueba se basa en la especificidad de los anticuerpos por un
antígeno, al cual se unen de manera similar a la llave con su cerradura. En esta
técnica inmunoenzimática, un antígeno inmovilizado en un soporte sólido
(generalmente placas de poliestireno) se detecta mediante un anticuerpo enlazado
a una enzima (ELISA directa) o un anticuerpo primario que reconoce al antígeno y
que a su vez es reconocido por un anticuerpo secundario que lleva enlazado la
enzima (ELISA indirecta). Al final, una reacción de color permite medir mediante
espectrofotometría la molécula de interés presente en la muestra que se está
analizando (63) (Fig 2.9).

GRUPO POLIFENOLES
19
Figura 2.9. Etapas de un ensayo de ELISA.
2.3.2 Reacción en cadena de la polimerasa con transcripción reversa (RT-
PCR)
La amplificación de los blancos del RNA mensajero (mRNA) son convertidos a
DNA complementario (cDNA) mediante la acción de una enzima llamada
transcriptasa reversa (RT), para luego ser amplificados por PCR.
La PCR es una técnica de amplificación de ácidos nucleicos; se basa en la
capacidad de la ADN polimerasa para copiar una cadena de DNA por
alargamiento de cadenas complementarias a partir de un cebador o “primer” con
sentido y antisentido. Su amplificación se da mediante varios ciclos en un
termociclador, los cuales se llevan a cabo mediante los tres siguientes pasos:
Desnaturalización del cDNA, en el cual se separan sus dobles cadenas a una alta
temperatura.
Unión de los “primers” o cebadores (alineación), en la cual al disminuir la
temperatura, se permite la unión de ellos a las secuencias complementarias del
cDNA.
Anticuerpo específico unido al soporte sólido
Adición del la muestra y unión del antígeno al anticuerpo
Adición del sengudo anticuerpo unido a la enzima
Adición del sustrato de la enzima
Desarrollo de color
:Anticuerpo
:Sustrato de la enzima
:Anticuerpo unido a una enzima

GRUPO POLIFENOLES
20
Extensión, la enzima ADN polimerasa extiende las secuencias entre los “primers”.
Al final de los ciclos el número de moléculas de cDNA se amplifica, después de
30-50 ciclos ocurre un crecimiento exponencial en el número total de copias de
cDNA sintetizadas (Fig 2.10) (64).
Transcriptasa reversa
mRNA
cDNA
Alineación
Desnaturalización
Extensión(ADN polimerasa)
3'
5'
5'
3'
"Primers" conantisentido
"Primers" consentido
5'
5'
5'
5' 3'
3'
3'
3'
5'
5'
5'
5'
3'3'
3'
3'
Después de XX ciclos
>1000000 de copias
Figura 2.10. Reacción en cadena de la polimerasa con transcriptasa reversa.

GRUPO POLIFENOLES
21
2.4 Anti-inflamatorios no esteroidales (AINES)
Los anti-inflamatorios no esteroidales (AINEs) son un amplio grupo de fármacos
empleados como anti-inflamatorios, analgésicos y antipiréticos. los AINEs tienen
diversas estructuras químicas (Fig. 2.11) sin embargo el mecanismo de acción se
basa en la inhibición de las enzimas COX-1 y/o COX-2, lo cual conlleva a una
inhibición en la producción de mediadores de la inflamación como prostaglandinas
y tromboxanos (65).
O
O
SO
O
H3C
Rofecoxib (inhibidor selectivo de COX-2)
OH
O
Ibuprofeno (AINEs)
O
O OH
O
Aspirina (AINEs)
Figura 2.11. Estructura química de algunos anti-inflamatorios.
Aunque los AINEs son muy efectivos para tratar la inflamación presentan algunos
efectos secundarios, producto de la inhibición de COX-1, la cual es expresada de
manera constitutiva en la mayoría de los tejidos y es la responsable de funciones
fisiológicas normales como el mantenimiento de la integridad de la mucosa
gástrica y la regulación del flujo sanguíneo renal (19), por lo tanto, al inhibirse la
COX-1, se generan gastropatías (36). Aunque existen inhibidores selectivos de
COX-2 que disminuyen la inflamación y son menos gastrolesivos, éstos pueden
generar efectos secundarios indeseables como problemas cardiovasculares (37).

GRUPO POLIFENOLES
22
3 SECCIÓN EXPERIMENTAL
3.1 Materiales, equipos y reactivos
3.1.1 Material vegetal y biológico
La especie Tabebuia chrysantha (JACQ.) G.Nicholson. se recolectó en Septiembre
de 2010 en el edificio de la Facultad de Ingeniería Industrial de la Universidad
Tecnológica de Pereira. Dicha planta ya había sido identificada en el Jardín
Botánico de la Universidad Tecnológica de Pereira y se confirmó en la Facultad de
Ciencias de la Universidad del Tolima.
Se empleó la línea celular RAW264.7 (monocitos - macrófagos de ratón
leucémico), la cual fue adquirida por el Laboratorio de Fisiología Celular e
Inmunología.
3.1.2 Equipos
Los equipos fueron suministrados por el Laboratorio de Fitoquímica y el
Laboratorio de Fisiología Celular e Inmunología.
Rotavaporador HEIDOLPH- 4003.
Balanza analítica OHAUS Adventurer AR2140.
Balanza electrónica OHAUS Explorer Pro EP4102.
Ultrasonido Fisher Scientific SF60H.
Horno BINDER ED-115.
Espectrofotómetro UV-Vis Shimadzu-UV-1700 Pharmaspec.
Jasco HPLC 2000 Plus. Sistema equipado con bomba de gradiente cuaternario
PU-2089 Plus, automuestreador inteligente AS-2059 Plus, horno para columna
CO-2065 Plus, columna analítica Ultra Aqueous C-18 Restek (2 µm tamaño de

GRUPO POLIFENOLES
23
partícula, 100 x 3.2 mm D.I), detector inteligente de arreglo de diodos MD-2015
Plus y LC Net II/ADC, controlado por el Software EZChrom Elite.
BIO-RAD. Molecular imager. Gel DOCTM XR+. With Image LabTM Software.
Model Universal Hood II.
Applied Biosystems Veriti 96well Thermal Cycler. Model 9902
Incubadora Thermo Electron Corporation Forma series II. Water Jacketed CO2
incubador. Hepa Class 100.
Centrifuge 5804R 15amp Version. Eppendorf.
Cámara de electroforesis. Bio-RAD y fuente de poder PowerPac HC.
Centrífuga Mini Spin plus. Eppendorf.
Cabina de flujo laminar C4 y Streamline.
Microscopio Nikon eclipse E100.
Balanza analítica OHAUS PioneerTM.
Microscopio invertido Leica.
Lector de placas Stat Fax 3200, AWARENESS Technology inc.
NanoDrop 2000c
3.1.3 Reactivos
Metanol grado industrial para la extracción (protokimica).
Hexano (Merck).
Cloroformo (Merck).

GRUPO POLIFENOLES
24
Acetato de etilo (J.T. Backer).
Butanol (Merck).
Acetona (Merck).
Metanol (Merck).
Reactivos marcha fitoquímica: Cloruro férrico, cloruro de sodio, ácido pícrico,
hidróxido de sodio, etanol, clorhidrato de hidroxilamina, hidróxido de potasio,
ácido clorhídrico, zin en polvo, ácido tricloroacetico, cloroamina T, ácido
sulfúrico, ácido acético, magnesio en polvo, alcohol amílico, anhídrido acético,
solución salina 0,9%, hidroxilamina, α-naftol, yoduro de mercurio II, yoduro de
potasio y subnitrato de bismuto.
Dimetilsulfóxido (Sigma).
RNeasy plus mini kit (Qiagen).
RNeasy mini kit (Qiagen).
BD OptEiA set mouse TNF (mono/mono) (BD Biosciences).
Griess reagent (modified) (Sigma).
3.2 Obtención de los extractos
Se tomaron 2 Kg de corteza interna de Tabebuia chrysantha previamente molida a
la cual se le realizó una extracción con metanol industrial y se aplicó ultrasonido
por 1 hora. Posteriormente se sometió a percolación durante 87 horas. El extracto
en metanol se filtró y se concentró a presión reducida.
Se tomaron 25 g del extracto en metanol y se disolvieron en 250 mL de metanol-
H2O (35% H2O). Posteriormente se realizaron extracciones sucesivas líquido-

GRUPO POLIFENOLES
25
líquido con hexano, cloroformo, acetato de etilo y butanol respectivamente (Fig
3.1).
2 Kg de Corteza interna
Percolar con metanol x 87 h
m=25,01 g
Concentrar el extracto en metanol
Extracto en n-Hexanom=0,7984 g
Metanol (35% H20)
n-Hexano
Extracto en Cloroformom=5,9546 g
Clorofórmo
Evaporar el metanol
Extracto en Acetato de Etilom=0,3940 g
Acetato de Etilo
Extracto en Aguam=10,6981 g
Butanol
Extracto en Butanolm=4,94721 g
Figura 3.1. Diagrama de extracción líquido – líquido de T. chrysantha.

GRUPO POLIFENOLES
26
3.3 Fraccionamiento del extracto en cloroformo
Del extracto en cloroformo de obtuvieron 5,9 g, de los cuales se tomó 1 g para el
fraccionamiento. Se utilizó una columna de vidrio con 80g de Silica gel 60 (0,063-
0,200 mm) (Merck). Como eluentes se usaron gradientes de 600 mL de n-hexano-
acetona y acetona-metanol en las proporciones descritas (Fig 3.2). Se
recolectaron 430 fracciones de 20 mL cada una, las cuales fueron reunidas de
acuerdo a sus perfiles cromatográficos por cromatografía en capa fina (TLC),
obteniéndose finalmente 21 fracciones, las cuales fueron concentradas a presión
reducida.
1 g de extracto en cloroformo
..........
F-1 F-2 F-3 F-5 F-6 F-7 F-8 F-9 F-10 F-11 F-19 F-20 F-21
VLecho= 200 mL
Vetapa = 600 mL
Gradientes n-Hexano-Acetona:
(10:0), (9,5:0,5), (9:1), (8,5:1,5),
(8:2), (7,5:2,5), (7:3), (6:4), (5:5),
(4:6).
Gradientes Acetona-Metanol:
(9:1), (6:4), (3:7), (1:9).
Figura 3.2. Fraccionamiento del extracto en cloroformo.

GRUPO POLIFENOLES
27
3.4 Perfiles cromatográficos por HPLC
Los perfiles cromatográficos de los extractos en n-hexano, cloroformo, acetato de
etilo, butanol y agua, se realizaron mediante cromatografía líquida de alta
eficiencia (HPLC), a una concentración de 2000 ppm y empleando una columna
de fase reversa Ultra Aqueous C-18 Restek de 3,2 mm de diámetro interno y 100
mm de largo.
Los extractos fueron eluidos con un gradiente de ácido fosfórico 0,05% (solvente
A): acetonitrilo (solvente B), variando las proporciones de éstos en un gradiente,
según se muestra en la Fig. 3.3. Los parámetros empleados en el equipo se
muestran en la Tabla 3.1:
Parámetros Condiciones
Detector PDA (200-701 nm)
Gradiente
Ácido fosfórico 0,05% : Acetonitrilo
90:10
68:32
45:55
68:32
90:10
Flujo 0,5 mL/min
Temperatura 35 °C
Tiempo de corrida 30,1 min
Volumen de inyección 20 µL
Tabla 3.1. Condiciones del HPLC-DAD para fracciones polares.

GRUPO POLIFENOLES
28
Figura 3.3. Gradiente de ácido fosfórico–acetonitrilo.
3.5 Marcha fitoquímica de la corteza interna de Tabebuia chrysantha
Se realizaron diferentes pruebas de caracterización fitoquímica al material vegetal
con el fin de conocer de manera preliminar los metabolitos secundarios presentes
en la corteza interna de T. chrysantha, siguiendo los procedimientos descritos en
materiales bibliográficos (66), (67) como se observa en la Fig. 3.4.
Para la identificación de los compuestos se emplearon diferentes pruebas, las
cuales se mencionan en la Tabla 3.2.

GRUPO POLIFENOLES
29
Compuesto Prueba
Taninos Gelatina-sal
FeCl3
Flavonoides Shinoda
Rosenhein
Cardiotónicos
Baljet
Molish
Liebermann-Burchard
Ácido tricloro acético
Saponinas Hemolisis de sangre
Espuma
Quinonas
Comportamiento frente a una base
Comportamiento frente a un ácido
Latonas
sesquiterpenicas Óleum
Alcaloides
Dragendorff
Valser
Bouchardat
Tabla 3.2. Pruebas de caracterización de compuestos químicos.

GRUPO POLIFENOLES
30
10 g Corteza interna
Lixiviar en etanol 95% y filtrar
FILTRADOA
RESIDUO(Descatar)
10 mL 5 mL 5 mL
Evaporar elsolvente
Evaporar elsolvente
Concentrar a lamitad delvolumen
EXTRACTO I EXTRACTO II EXTRACTO III
Esteroides,flavonoides,quinonas,saponinas,taninos.
Lactonas terpenicas,cumarinas,cardiotonicos.
Alcaloides.
Realizar pruebas para
Realizar pruebas para
Realizar pruebas para
Figura 3.4. Marcha fitoquímica de la corteza interna de T. chrysantha.
3.6 Ensayos biológicos
3.6.1 Test de viabilidad celular
En una placa de 96 pozos se incubó la línea celular de macrófagos murinos
RAW264.7 (5x104 células/pozo) durante 24 horas en medio de cultivo, el cual

GRUPO POLIFENOLES
31
consistió en Dulbecco's Modified Eagle Medium (DMEM) con Glutamax II
(GIBCO/BRL, USA), 10% de suero bovino fetal inactivado por calor, antibióticos
(200 μg/mL penicilina, 200 μg/mL estreptomicina y 400 μg/mL neomicina), 10
mg/mL anfotericina B, 1 mM piruvato de sodio y 0,05 mM de 2-β-mercaptoetanol.
Posteriormente se adicionaron los extractos a diferentes concentraciones (0,625-
2,5 µg/mL) y se incubaron las células durante 24 horas. Después del tratamiento,
se descartó el medio de cultivo y se adicionaron 200 µL de una solución de MTT (5
mg/mL) a cada pozo. Se incubaron las placas durante 4 horas a 37 ⁰C bajo una
atmósfera de 5% de CO2. Luego de descartar el medio, se agregaron 100 μL de
isopropanol 0,04 N en HCl con el fin de solubilizar el formazán generado. La
densidad óptica se midió a 492 nm en un lector de placas (Stat Fax 3200,
Awareness technology Inc., USA).
3.6.2 Determinación de la producción de PGE2 y TNF-α
En una placa de 96 pozos se incubaron macrófagos RAW264.7 (1x106
células/pozo) durante 24 horas y durante las últimas 22 horas de cultivo se
adicionó una solución de ácido acetilsalicílico (50 µM), con el fin de inactivar la
COX-1 endógena. Luego de lavar con PBS y agregar medio de cultivo fresco, se
estimularon los macrófagos con LPS (5 µg/mL durante 12 h), con el fin de detectar
los niveles de PGE2/TNF-α. La concentración de LPS y los tiempos de
estimulación se seleccionaron basados en experimentos cinéticos previos. Las
células se estimularon con LPS en presencia o ausencia de los extractos
vegetales (2,5 µg/mL). Como control, se empleó medio de cultivo sin la presencia
de LPS. En todos los casos, la concentración final de DMO en el cultivo fue menor
del 0,2%. El nivel de endotoxina en los extractos se evaluó empleando el kit E-
toxate (Sigma). Todas las muestras fueron negativas para la presencia de
endotoxina (límite de detección 0,05 – 0,1 EU/mL).
La concentración de PGE2 en el medio de cultivo se determinó usando un kit
comercial de Cayman Chemical Co. (Ann Arbor, Mi, USA), siguiendo las

GRUPO POLIFENOLES
32
instrucciones del fabricante. El porcentaje de inhibición en la producción de la
PGE2 se expresó como [1 – (PGE2 en la muestra/ PGE2 en el blanco – PGE2 en el
control)] x 100. La concentración de TNF-α se determinó empleando kits de ELISA
comerciales (OptEIA ELISA Set, BD Biosciences, USA), Siguiendo las
indicaciones del fabricante. El porcentaje de inhibición en la producción de la TNF-
se expresó como [1 – (TNF-α en la muestra/TNF-α en el blanco – TNF-α en el
control)] x 100.
3.6.3 Determinación de la producción de NO
La estimulación de los macrófagos se realizó de manera similar a la realizada para
detectar los niveles de PGE2 y TNF-α. A diferencia de dicho experimento, se
estimuló a los macrófagos murinos con 10 µg/mL de LPS durante 18 h. La
concentración de NO en el medio de cultivo se determinó mezclando 100 µL del
sobrenadante de cultivo con 100 µL del reactivo de Griess (50 µL 1% sulfanilamida
en 5% H3PO4 y 50 µL 0.1% N-1-naptiletilendiamina) durante 5 min. Se determinó
la absorbancia a 550 nm. Se empleó medio de cultivo fresco como blanco en
todos los experimentos. La cantidad de nitrito en cada muestra se calculó usando
una curva standard preparada con nitrito de sodio (NaNO2). El porcentaje de
inhibición en la producción de NO se expresó como [1 – (NO en la muestra/NO en
el blanco – NO en el control)] x 100.
3.6.4 Extracción de RNA y análisis por medio de RT-PCR (transcripción
reversa reacción en cadena de la polimerasa)
Se extrajo el RNA de las células RAW264.7 tratadas con los extractos obtenidos
de la corteza interna de T. chrysantha y estimuladas con LPS, utilizando el Kit
RNeasy mini kit y siguiendo las instrucciones del fabricante. Dicho RNA se
almacenó a – 80 °C hasta su uso.
La evaluación de la expresión de los genes COX-2, TNF-α e iNOS se realizó
mediante una RT-PCR empleando el Kit GeneAmp Gold RT-PCR (Applied

GRUPO POLIFENOLES
33
Biosystems), siguiendo las instrucciones del fabricante. Los primers utilizados se
muestran en el ANEXO 1.
3.6.5 Cuantificación de RNA
Para determinar la pureza y concentración del RNA extraído se utilizó un
espectrofotómetro NanoDrop 2000/2000c, utilizando como blanco agua libre de
RNasas. Los resultados fueron analizados mediante el software Nanodrop
2000/2000c versión 1.4.2.
3.6.6 Electroforesis de los productos de RT-PCR
Los productos de RT-PCR se sometieron a electroforesis en un gel de agarosa al
2%, utilizando como marcador de peso molecular Hyperladder II y como agente
intercalante SyBR-safe. Se aplicaron 80 voltios durante 90 minutos. El gel se
reveló en un fotodocumentador BIO-RAD. Molecular Imager. Gel DOCTM XR+.
With Image LabTM Software.
3.7 Fraccionamiento de la fracción F-16 del extracto en cloroformo
De la fracción F-16 se tomaron aproximadamente 200 mg para el fraccionamiento.
Se utilizó una columna de vidrio en fase reversa empacada con octadecilsilano
(Aldrich). Como eluentes se usaron 400 mL de metanol, 400 mL de acetona y
finalmente 400 mL de acetato de etilo (Fig 3.5). Se recolectaron 9 fracciones, las
cuales fueron concentradas a presión reducida.

GRUPO POLIFENOLES
34
200 mg de fracción F16
F-1 F-2 F-3 F-4 F-5 F-6 F-7 F-8 F-9
Vetapa = 400 mL
Eluentes:
Metanol
Acetona
Acetato de etilo
Figura 3.5. Fraccionamiento de la fracción F-16 del extracto en cloroformo
3.8 Purificación de compuestos de la F-16-3 de la fracción F-16
De la fracción F-16-3 se tomaron aproximadamente 80 mg para su separación,
mediante cromatografía preparativa. Se utilizó una placa de vidrio de silica gel 60
F254 de 20x20 cm con 1 mm de espesor (Merck). Como eluente se utilizó una
mezcla de 100 mL de diclorometano-acetona (9:1). Se obtuvieron 6 fracciones, las
cuales fueron concentradas a presión reducida.
3.9 Espectroscopia de resonancia magnética nuclear (RMN)
Los espectros de RMN de 1H y 13C de la fracción F-16-3-b, fueron obtenidos
mediante un equipo de resonancia magnética nuclear (Bruker Avance III 400
MHz). La fracción fue disuelta en 0,7 mL de una mezcla de acetona- agua (5:2).

GRUPO POLIFENOLES
35
3.10 Análisis estadístico
Todos los experimentos de estimulación se realizaron tres veces por duplicado. Se
expresan los resultados como Media ± SEM. Dado que no fue posible determinar
la normalidad de los datos debido al tamaño de la muestra, se realizaron
comparaciones mediante la prueba de Kruskal-Wallis con una prueba “Pos – hoc”
correspondiente al test de comparaciones múltiples de Dunn´s, Se consideraron
estadísticamente significativos valores de p ≤ 0,05. El análisis estadístico se
realizó con el programa GraphPad Prism versión 6,0 (GraphPad Software, San
Diego, CA, USA).

GRUPO POLIFENOLES
36
4 RESULTADOS Y DISCUSIÓN
4.1 Rendimiento de extracción
Se obtuvieron 132,9 g de extracto en metanol de la corteza interna de T.
chrysantha con un rendimiento de extracción de 6,6% con respecto al material
vegetal seco y molido.
El porcentaje de rendimiento de extracción en todo el proceso fue de 91 % y los
rendimientos obtenidos para las extracciones sucesivas del extracto en metanol de
corteza interna de T. chrysantha se muestran en la Fig. 4.1.
n-Hex
ano
Clo
rofo
rmo
AcO
Et
BuO
H
Agua
0
10
20
30
40
50
% d
e r
en
dim
ien
to
Figura 4.1. Rendimiento de extracción líquido-líquido del extracto en
metanol.
Se puede observar que el extracto en n-hexano y acetato de etilo tienen un
rendimiento muy bajo (3,2 y 1,6%, respectivamente), mientras que los extractos en
cloroformo, butanol y agua presentan un alto rendimiento (23,8, 19,8 y 42,8%,
respectivamente). Lo anterior indica que la mayoría de los compuestos están
presentes en los extractos de mayor polaridad (Fig. 4.1).

GRUPO POLIFENOLES
37
4.2 Perfiles cromatográficos por HPLC de los extractos de corteza interna
de T. chrysantha
A todos los extractos de la corteza interna de T.chrysantha se les realizó un perfil
cromatográfico por HPLC. Los cromatogramas obtenidos para los extractos en n-
hexano, butanol y agua (VER ANEXO 2, 3 Y 4) no brindan información suficiente
para hacer un análisis sobre la naturaleza de los compuestos que contienen, a
diferencia de los perfiles cromatográficos de los extractos en cloroformo (Fig. 4.2)
y acetato de etilo (Fig. 4.3) que se muestran a continuación.
Figura 4.2. Perfiles cromatográficos del extracto en cloroformo. (A)
Cromatograma, (B) Espectro UV/VIS del pico 4 y (C) Espectro UV/VIS del pico
7.

GRUPO POLIFENOLES
38
Figura 4.3. Perfiles cromatográficos del extracto en acetato de etilo. (A)
Cromatograma, (B) Espectro UV/VIS del pico 2 y (C) Espectro UV/VIS del pico
7.
Se puede observar que los picos 4 y 7 del cromatograma del extracto en
cloroformo y los picos 2 y 7 del extracto en acetato de etilo absorben intensamente
entre 240-290 nm. Este tipo de absorbancia es característica de las naftoquinonas,
antraquinonas, benzoquinonas y homólogos (67). Se compararon los espectros de
los extractos antes mencionados con espectros reportados para compuestos de
este tipo (Fig. 4.4) (68), observándose gran similitud entre estos. Teniendo en

GRUPO POLIFENOLES
39
cuenta lo anterior y los resultados obtenidos en la marcha fitoquímica, se puede
inferir la posible presencia de compuestos de la familia quinona.
Figura 4.4. Espectro UV/VIS de la 1,4-benzoquinona.
4.3 Marcha fitoquímica
La caracterización preliminar de los compuestos químicos presentes en la corteza
de T chrysantha se realizó mediante una marcha fitoquímica, cuyos resultados se
muestran en la Tabla 4.1.

GRUPO POLIFENOLES
40
Compuesto Prueba Pruebas de tubos TLC
Taninos Gelatina-sal - *
FeCl3 - -
Flavonoides
Shinoda - *
Rosenhein + *
Cardiotónicos
Baljet + *
Molish + *
Liebermann-Burchard + *
Ácido tricloro acético * +
Saponinas
Hemolisis de sangre + *
Espuma - *
Quinonas
Comportamiento frente a una base + *
Comportamiento frente a un ácido + *
Latonas
sesquiterpenicas Óleum * +
Alcaloides
Dragendorff - *
Valser - *
Bouchardat - *
Tabla 4.1. Resultados de la marcha fitoquímica del extracto de corteza
interna de T. Chrysantha.
Convenciones: *: No se realizó.
+: Prueba positivo
-: Prueba negativa

GRUPO POLIFENOLES
41
Con la caracterización preliminar se puede inferir que la corteza interna de T.
chrysantha, presenta glicósidos cardiotónicos, quinonas y lactonas
sesquiterpenicas.
4.4 Ensayo de viabilidad celular
Previo a la evaluación de la actividad anti-inflamatoria se determinó la viabilidad
celular de los macrófagos murinos RAW264.7 frente a los extractos de corteza
interna de T. chrysantha a tres concentraciones expresadas en [µg/mL] (VER
ANEXO 5). A continuación se presentan los resultados (Fig. 4.5).
n-hex
ano [2
,5]
n-hex
ano [1
,3]
n-hex
ano [0
,6]
Clo
rofo
rmo [2
,3]
Clo
rofo
rmo [1
,2]
Clo
rofo
rmo [0
,6]
AcO
Et [2,
4]
AcO
Et [1,
2]
AcO
Et [0,
6]
BuO
H [2
,1]
BuO
H [1
,1]
BuO
H [0
,5]
Agua
[2,2
]
Agua
[1,1
]
Agua
[0,6
]
Contr
ol (+)
Contr
ol (-)
0
50
100
150
200
% V
iab
ilid
ad
Figura 4.5. Viabilidad celular de los macrófagos murinos RAW264.7 frente a
los extractos de corteza interna de T. chrysantha.
Se logró determinar que todos los extractos presentan una viabilidad celular
superior al 70% con respecto al control positivo a concentraciones menores o
iguales a 2,5 µg/mL, excepto el extracto en butanol que presenta una mayor
viabilidad (89%) a una concentración menor o igual a 1,1 µg/mL (Fig 4.5).
Teniendo en cuenta lo anterior se seleccionó la concentración más alta de los

GRUPO POLIFENOLES
42
extractos para los bioensayos, a diferencia del extracto en butanol, para el cual se
utilizó la concentración de 1,1 µg/mL.
4.5 Actividad anti-inflamatoria in vitro de los extractos de T.chrysantha.
La línea celular de macrófagos murinos RAW264.7 estimulados con LPS generan
una serie de mediadores de la inflamación tales como PGE2, TNF-α y NO (25). En
estos ensayos se determinó la capacidad de los extractos de la corteza interna de
T. chrysantha para inhibir la producción de dichos mediadores.
Para la determinación de la inhibición en la producción de TNF-α y NO se usaron
dos controles; LPS (+) y LPS (-) que permiten ver la máxima y mínima producción
del mediador respectivamente. Para PGE2 se utilizaron dos controles adicionales a
los mencionados anteriormente, Asp-LPS (+), en el cual inhibe selectivamente la
enzima COX-1 y Asp-DUP-LPS (+), en el cual se inhiben tanto COX-1 como COX-
2. A continuación se muestran los resultados.
4.5.1 Inhibición en la producción de PGE2
Los macrófagos murinos RAW264.7 estimulados con LPS en presencia de los
extractos en n-hexano, cloroformo, acetato de etilo butanol y agua inhibieron la
producción de PGE2 en un rango entre el 5% y el 47% (VER ANEXO 6). El mayor
efecto inhibitorio se observó con el extracto en cloroformo (47%), mientras que el
extracto en butanol mostró la menor inhibición (5%). (Fig 4.6). Sin embargo, no se
encontraron diferencias significativas al comparar con el control de inhibición Asp-
DUP-LPS (+).

GRUPO POLIFENOLES
43
n-Hex
ano
Clo
rofo
rmo
AcO
Et
BuO
H
Agua
LPS (-
)
LPS (+
)
Asp
, LPS (+
)
Asp
, Dup, L
PS (+
)
0
50
100
150
% I
nh
ibic
ión
de P
GE
2
Figura 4.6. Efecto inhibitorio de los extractos de la corteza interna de T.
chrysantha sobre la producción de PGE2 en macrófagos murinos
estimulados con LPS.
4.5.2 Inhibición en la producción de TNF-α
La estimulación de los macrófagos murinos RAW264.7 con LPS indujo a la
producción de citocinas pro-inflamatorias como el TNF-α. Los extractos en n-
hexano, cloroformo, acetato de etilo y agua mostraron los niveles más altos de
inhibición en la producción de TNF-α, destacándose el extracto en n-hexano, con
una inhibición del 112 %, mientras que el extracto en butanol mostró la menor
inhibición con 5% (VER ANEXO 7). Se observaron diferencias estadísticamente
significativas para los extractos en n-hexano (p=0,030), cloroformo (p=0,004) y
acetato de etilo (p=0,038) comparados con el control LPS (+) (Fig. 4.7).

GRUPO POLIFENOLES
44
n-Hex
ano
Clo
rofo
rmo
AcO
Et
BuO
H
Agua
LPS (-
)
LPS(+
)
0
50
100
150
*P=0,030
**P=0,004
*P=0,038
% I
nh
ibic
ión
de T
NF
-
Figura 4.7. Efecto inhibitorio de los extractos de la corteza interna de T.
chrysantha sobre la producción de TNF-α en macrófagos murinos
estimulados con LPS.
4.5.3 Inhibición en la producción de óxido nítrico (NO)
La producción de NO por macrófagos murinos RAW 264.7 estimulados con LPS
se evaluó midiendo la concentración de nitritos presentes en el medio de cultivo,
mediante una reacción de coloración empleando el reactivo de Griess. Los
macrófagos estimulados en presencia de los extractos en n-hexano, acetato de
etilo y agua aumentan la producción de NO en un porcentaje superior al 100%
comparados con el control LPS (+). El extracto en cloroformo promueve la
producción en un 13% mientras que por el contrario, el extracto en butanol inhibe
la producción de NO en un 60%. (VER ANEXO 8)
Se observó una diferencia estadísticamente significativa (p=0,011) en la
producción de NO entre el extracto en n-hexano comparado con el control LPS (-),
para los demás extractos no se observaron diferencias significativas (Fig 4.8).

GRUPO POLIFENOLES
45
n-Hex
ano
Clo
rofo
rmo
AcO
Et
BuO
H
Agua
LPS (-
)
LPS (+
)-600
-400
-200
0
200*P=0,011
% I
nh
ibic
ión
de N
O
Figura 4.8. Efecto inhibitorio de los extractos de la corteza interna de T.
chrysantha sobre la producción de NO en macrófagos murinos estimulados
con LPS.
4.6 CUANTIFICACIÓN DE mRNA EN LA LINEA CELULAR RAW264.7
ESTIMULADAS CON LPS
La normalización de las concentraciones del mRNA de los macrofagos RAW264.7
estimulados con LPS y tratados con los extractos de corteza interna de T.
chrysantha, se llevó a cabo mediante técnicas espectrofotométricas, las lecturas
se realizaron utilizando como blanco el RNasa Free Water del RNeasy Plus mini
kit. Los resultados se muestran en la Tabla 4.2.

GRUPO POLIFENOLES
46
Muestra Concentración
(µg/µl)
A260
(nm)
A280
(nm) 260/280
n-Hexano 0,233 5,827 2,791 2,090
Cloroformo 0,238 5,949 2,834 2,100
Acetato de etilo 0,247 6,185 2,946 2,097
Butanol 0,264 6,612 3,140 2,110
Agua 0,241 6,034 2,881 2,087
LPS (-) 0,190 4,541 2,158 2,113
LPS (+) 0,219 5,634 2,668 2,110
Asp, LPS (+) 0,283 7,168 3,405 2,107
Asp, DUP, LPS (+) 0,212 6,627 3,156 2,100
Tabla 4.2. Cuantificación de mRNA de los macrofagos RAW264.7
estimulados con LPS.
Mediante la cuantificación de mRNA se estableció la cantidad a emplear en la
mezcla de reacción de la RT-PCR.
Debido a que las bases púricas y pirimidínicas del DNA absorben a 260 nm y las
proteínas a 280 nm, la relación entre estas dos absorbancias (260/280) se utiliza
para evaluar la pureza del RNA. Dicha relación debe estar en el rango entre 1,8 -
2,0 (69). Dado que todas las muestras de mRNA obtenidas a partir de macrofagos
RAW264.7 estimulados con LPS y tratados con los extractos de corteza interna de
T. chrysantha, se encuentran en dicho rango, podemos decir que dicho mRNA
está libre de contaminantes como proteínas o fenoles (70).
4.7 Efecto de los extractos sobre la expresión de mRNA de COX-2, TNFα e
iNOS en macrófagos murinos RAW264.7 estimulados con LPS.
Para establecer si hay una relación entre la inhibición de la producción de los
mediadores de inflamación (PGE2, TNF-α y NO) y la regulación de las expresiones

GRUPO POLIFENOLES
47
de COX-2, TNF-α e iNOS, se realizó una RT-PCR a partir del mRNA obtenido de
los macrófagos RAW 264.7 estimulados con LPS, usando como gen control β-
actina y un marcador de peso molecular Hyperladder II (100-2000 pb). En Fig. 4.9
se muestran los resultados obtenidos, expresados en forma de intensidad relativa
con respecto al gen control.
n-Hex
ano
Clo
rofo
rmo
Ace
tato
de
etilo
Buta
nol
Agua
LPS (-
)
LPS (+
)
Asp
, LPS (+
)
Asp
, Dup, L
PS (+
)
0
1
2
3
COX-2
TNF-a
iNOs
Niv
ele
s r
ela
tivo
s d
e m
RN
A
Figura 4.9. Efecto inhibitorio de las expresiones de mRNA de COX-2, TNF-α e
iNOS sobre macrófagos murinos RAW264.7 estimulados con LPS.
Se observa que la línea celular de macrófagos murinos RAW264.7 presenta bajos
niveles relativos de expresión de los genes COX-2, TNF-α e iNOS, cuando no han
sido estimulados, a diferencia de las células que han sido tratadas con LPS (71),
en las cuales se incrementa la expresión de dichos genes.
Con relación a la expresión de COX-2, los extractos en n-hexano y cloroformo
inducen una baja expresión de dicho gen, mientras que los extractos en acetato de
etilo, butanol y agua promueven la expresión del gen, al comparar con el control
de estimulación LPS (+).

GRUPO POLIFENOLES
48
Los extractos en n-hexano, cloroformo, butanol y agua incrementaron la expresión
de TNF-a comparado con el control de estimulación LPS (+), mientras que el
extracto en acetato de etilo disminuyó su expresión.
Los extractos en n-hexano y acetato de etilo disminuyeron la expresión de iNOS,
mientras que los extractos en cloroformo, butanol y agua no alteraron su expresión
al comparar con el control de estimulación LPS (+).
No se encontraron diferencias estadísticamente significativas en los patrones de
expresión de los genes analizados.
Los macrófagos murinos RAW 264.7 estimulados con LPS producen una serie de
mediadores de la inflamación tales como la PGE2, implicada en la vasodilatación y
el NO, involucrado en los procesos de vasodilatación y reducción de la agregación
y adhesión plaquetaria. Los macrófagos estimulados con LPS también secretan
otro tipo de sustancias como las citocinas pro-inflamatorias, entre las que se
encuentran IL-6, IL-8, IL-1β y TNF-α (22). En el presente estudio se evaluó el
efecto de los extractos obtenidos de la corteza interna de T.chrysantha sobre la
producción de PGE2, NO y TNF-α y la expresión de los genes que codifican por
las enzimas responsables de la producción de PGE2 y NO (COX-2 e iNOS,
respectivamente).
Todos los extractos de corteza interna de T.chrysantha inhiben la producción de
PGE2, destacándose el extracto en cloroformo, el cual presenta la mayor inhibición
(47%), al evaluar la expresión de COX-2 se puede observar que los extractos en
n-hexano y cloroformo disminuyen la expresión del gen, indicando que esta
disminución se encuentra directamente relacionada con la inhibición en la
producción de PGE2. Por el contrario, los extractos en acetato de etilo y agua
mostraron un incremento en la expresión de COX-2 e inhibición en la producción
de PGE2, indicando que estos extractos tienen efecto directo sobre la actividad
catalítica de la enzima COX-2. Esto se confirma al emplear el control de DUP 697,
el cual es un inhibidor selectivo de COX-2 que inhibe la actividad de la enzima y
para compensar este efecto la célula incrementa la expresión de COX-2 (72).

GRUPO POLIFENOLES
49
El extracto en butanol inhibe la producción de NO en un 60%, pero la expresión de
de iNOS no disminuye lo que sugiere que el mecanismo responsable de la
inhibición de NO es un efecto directo de estos compuestos sobre la actividad de la
enzima, a diferencia de los extractos en n- hexano, cloroformo, acetato de etilo y
agua que promueven la producción de NO.
Todos los extractos de corteza interna inhiben la producción de TNF-α comparado
con el estímulo de LPS (+) en porcentajes superiores al 70%, presentando una
mayor inhibición el extracto en n-hexano (116%).
Éstos resultados indican que la corteza interna de T. chrysantha tiene un efecto
inhibitorio in vitro sobre la producción de mediadores de la inflamación en
macrófagos murinos, dando soporte a los usos etnomédicos reportados en
Sudamérica.
El extracto en cloroformo se puede considerar como el más promisorio de todos
los evaluados, dado que inhibe la producción de PGE2 y TNF- y adicionalmente
inhibe la expresión de la enzima COX-2. Por lo tanto, dicho extracto podría
convertirse en una fuente de compuestos o metabolitos secundarios que pueden
llegar a ser utilizados como anti-inflamatorios. Estos resultados llevaron al
fraccionamiento del extracto en cloroformo, con el fin de identificar los compuestos
responsables de dicha actividad.
4.8 Rendimiento del fraccionamiento del extracto en cloroformo.
Dado que el extracto en cloroformo fue el más promisorio en la actividad biológica,
se continuó con el fraccionamiento de éste. El porcentaje de rendimiento del
fraccionamiento en columna fue del 97,76%. A continuación se presenta el
rendimiento de las fracciones obtenidas de dicho extracto en la Tabla 4.3.

GRUPO POLIFENOLES
50
Fracción Tc-F1
Tc-F2
Tc-F3
Tc-F14
Tc-F5
Tc-F6
Tc-F7
Tc-F8
Tc-F9
Tc-F10
Tc-F11
Rendimiento (%)
0,67 0,69 0,80 1,01 0,90 0,53 0,75 1,17 0,30 0,47 1,96
Fracción Tc-F12
Tc-F13
Tc-F14
Tc-F15
Tc-F16
Tc-F17
Tc-F18
Tc-F19
Tc-F20
Tc-F21
Rendimiento (%)
49,88 7,33 1,94 3,64 5,91 14,05 0,90 2,82 1,79 0,28
Tabla 4.3. Rendimiento del fraccionamiento del extracto en cloroformo
Se puede observar que el porcentaje de rendimiento de las fracciones (Tabla 4.3)
Tc-F1 a Tc-F11 es muy bajo comparado con el resto de las fracciones, indicando
que estas primeras son de carácter apolar, por lo tanto, la mayoría de los
compuestos presentes en el extracto en cloroformo son de alta polaridad.
4.9 Viabilidad celular de las fracciones del extracto en cloroformo
Posterior al fraccionamiento del extracto en cloroformo se realizó el ensayo de
viabilidad celular para determinar qué fracciones podían emplearse en los
posteriores ensayos de actividad anti-inflamatoria, con el fin de identificar los
compuestos responsables de dicha actividad. Los resultados obtenidos se
muestran en la Tabla 4.4.

GRUPO POLIFENOLES
51
Fracciones
Muestra Concentración
(μg/mL) % de
viabilidad Muestra
Concentración (μg/mL)
% de viabilidad
Tc-F1 1 98,1
Tc-F12
2 72,2
0,5 110,5 1 103
Tc-F3
2 77,4 0,5 120,4
1 98,2
Tc-F13
2 97,7
0,5 121,9 1 89,2
Tc-F4
2 18,7 0,5 117,4
1 56,8
Tc-F14
2 80,8
0,5 98,3 1 85
Tc-F5
2 73,9 0,5 109
1 85,7
Tc-F15
2 89,5
0,5 100,2 1 77,4
Tc-F6
2 78,7 0,5 105,6
1 70,2
Tc-F16
2 80,8
0,5 118,2 1 102,4
Tc-F7
2 62,3 0,5 118,3
1 98,4
Tc-F17
2 85,3
0,5 122,9 1 86,3
Tc-F8
2 67,2 0,5 103,8
1 62,9
Tc-F18
2 100,6
0,5 101,7 1 78,5
Tc-F9 1 97,6 0,5 87,6
0,5 103,4
Tc-F19
2 99,8
Tc-F10
2 77,8 1 94
1 88,5 0,5 117,7
0,5 113,5
Tc-F11
2 76,6
1 69,4
0,5 107,4
Controles
Control % de
viabilidad
Control positivo con células 100
Control negativo sin células 0
Tabla 4.4. Viabilidad celular de los macrófagos RAW264.7 frente a las
fracciones del extracto en cloroformo de T.chrysantha.

GRUPO POLIFENOLES
52
La mayoría de las fracciones del extracto en cloroformo de T. chrysantha
presentan una viabilidad celular superior al 80% a una concentración menor o
igual a 2 μg/mL con respecto al control positivo. Las fracciones Tc-F1, Tc-F7y Tc-
F9 mostraron una viabilidad superior al 80% a concentraciones menores o iguales
a 1 μg/mL, mientras que las fracciones Tc-F4 y Tc-F8 mostraron porcentajes de
viabilidad inferiores al 70% y por tanto, se descartaron para realizar los ensayos
de actividad anti-inflamatoria.
Teniendo en cuenta estos resultados y la poca cantidad de masa de las fracciones
se decidió unir algunas, (Tc-F13+Tc-F14) que se nombro como F13 y (Tc-F16+Tc-
F17+Tc-F18+Tc-F19) la cual se nombro como F16, debido a que la fracción Tc-
F12 tenía suficiente masa no se unió con otras fracciones, pero se nombro
nuevamente como F12. Finalmente para posteriores ensayos se usaron las tres
fracciones F12, F13 y F16.
4.10 Efecto de las fracciones del extracto en cloroformo de T. chrysantha
sobre la producción de PGE2, TNF-α y NO
Los macrófagos murinos RAW264.7 fueron estimulados con LPS en presencia de
las fracciones F12, F13 y F16 del extracto en cloroformo de T. chrysantha. Se
observó que las fracciones F13 y F16, fueron las que presentaron mayor inhibición
en la producción de PGE2 (35% y 40% respectivamente), NO (67% y 25%
respectivamente) y TNF-α (49% y 44% respectivamente). La fracción F12 inhibió
la producción de PGE2 (46,5%) y TNF-α (35%), sin embargo, estimuló la
producción de NO (Fig 4.10). Se observaron diferencias estadísticamente
significativas para la fracción F12 comparada con el control LPS (-) para la
inhibición en la producción de TNF-α (p=0,0450) y en la producción de NO
(p=0,0010).
Las fracciones F13 y F16 del extracto en cloroformo presentan los mayores
porcentajes de inhibición, por lo tanto se puede inferir que los metabolitos

GRUPO POLIFENOLES
53
responsables de la actividad anti-inflamatoria del extracto se encuentran
concentrados en estas fracciones.
F12 F13 F16
LPS (-
)
LPS (+
)
LPS+A
SP
ASP
+LPS+D
UP
0
50
100
150
A
% I
nh
ibic
ión
de P
GE
2
F12 F13 F16
LPS(-)
LPS(+
)0
50
100
150
*p=0,0450
B
% I
nh
ibic
ión
de T
NF
-
F12 F13 F16
LPS(-)
LPS(+
)-300
-200
-100
0
100
200 ***p=0,0010
C
% I
nh
ibic
ión
de N
O
Figura 4.10. Efecto inhibitorio de las fracciones del extracto en cloroformo de
la corteza interna de T. chrysantha sobre la producción de PGE2 (A), TNF-α
(B) y NO (C) en macrófagos murinos estimulados con LPS.

GRUPO POLIFENOLES
54
4.11 Rendimiento del fraccionamiento de la fracción F16 del extracto en
cloroformo
Dado que la fracción F16 presentó actividad anti-inflamatoria, se continúo con su
fraccionamiento, en la tabla 4.5 se muestra el rendimiento de las fracciones
obtenidas.
Fracción % de rendimiento
F16-1 …
F16-2 0,5
F16-3 46,8
F16-4 13,3
F16-5 8,1
F16-6 1,5
F16-7 0,6
F16-8 0,7
F16-9 0,3
Tabla 4.5. Rendimiento del fraccionamiento de la fracción F16.
Se observa que la mayor cantidad de compuestos son de carácter polar debido a
que hay gran cantidad de masa en las primeras fracciones, esto se debe al tipo de
columna empleada (octadecilsilano), por tanto los compuestos polares son los
primeros en eluir.
4.12 Purificación de compuestos de la fracción F16-3
Dado que la fracción F16-3 presentó la mayor cantidad de masa, se decidió
purificar mediante cromatografía preparativa, se obtuvieron 6 fracciones de las
cuales, la fracción F16-3-b mostró ser la menos compleja mediante espectros de
Resonancia Magnética Nuclear RMN 1H (Fig. 4.11) y 13C (Fig. 4.12) en

GRUPO POLIFENOLES
55
comparación con las otras fracciones que mostraron tener altas cantidades de
grasas vegetales.
Figura 4.11. Espectro de RMN de 1H de la fracción F16-3-b

GRUPO POLIFENOLES
56
Figura 4.12. Espectro de RMN de 13C de la fracción F16-3-b
En el espectro de RMN de 1H (Fig. 4.11) se observan en la región de aromáticos
dos dobletes con acoplamiento orto J:8,4 Hz entre (7,59-7,56) ppm, un doblete con
acoplamiento meta J:1,8 Hz entre (7,46-7,45) ppm, un doblete entre (6,99-6,97)
ppm con acoplamiento orto J:8,7 Hz, un multiplete (3,81-3,79) ppm
correspondiente a hidrógenos provenientes de grupos metoxilos; de otro lado en el
espectro de RMN de 13C (Fig. 4.12) se observan 9 carbonos, dentro de los cuales
se encuentran 3 carbonos cuaternarios aromáticos entre (153,24-148,75) ppm, 3
carbonos terciarios aromáticos entre (112,53-107,43) ppm y 3 carbonos primarios
correspondientes a grupos metoxilos entre (56,29-55,83) ppm. Con base en al
análisis anterior, se puede inferir que se trata de un sistema aromático tri-
sustituido que podría tener como sustituyentes grupos metoxilos (Fig.4.13), sin
embargo en los espectros de 1H y 13C se observan señales en 7,46 ppm y 124,33

GRUPO POLIFENOLES
57
ppm respectivamente, las cuales corresponden a otros compuestos o impurezas y
no permiten la clara elucidación de la estructura.
H
H
H
R1
R3
R2
R1:OCH3
R3:OCH3
R2:OCH3
Figura 4.13.Posible estructura de compuesto mayoritario de la fracción F16-
3-b
Al comparar con la literatura se han reportado compuestos derivados del ácido
benzoico aislados de la corteza de otras Tabebuias como el ácido veratrico
(Fig.4.13) aislado de Tabebuia heptaphylla (73), que tienen similitud con el
sistema aromático propuesto para el compuesto mayoritario presente en la
fracción F16-3-b.
O
OH
O
O
7.786,93
7,608,6
3,94
3,98
Ácido 3,4-dimetoxibenzóico
(Ácido veratrico)
Figura 4.14. Espectro de RMN de 1H del ácido veratrico.

GRUPO POLIFENOLES
58
5 CONCLUSIONES
Se puede observar que los compuestos del extracto en metanol de corteza
interna de T. chrysantha son principalmente de carácter polar, debido a que el
mayor porcentaje de rendimiento de los extractos son para el extracto en agua.
La viabilidad celular de los macrófagos murinos tratados con los extractos de
corteza interna de T. chrysantha, es superior al 90% con respecto al control.
La mayoría de los extractos mostraron una inhibición superior al 20% en la
producción de PGE2, excepto el extracto en butanol, el cual mostró una
inhibición menor al 10%.
La mayoría de los extractos inhibieron completamente la producción de TNF-α,
excepto el extracto en butanol el cual mostró una inhibición inferior al 90%.
En la determinación de la producción de óxido nítrico (NO) se pudo determinar
que los extractos en n-hexano, cloroformo, acetato de etilo y agua promueven
la producción de este mediador mientras que el extracto en butanol inhibió su
producción.
De todos los extractos evaluados, el extracto en cloroformo fue el más
promisorio por su actividad anti-inflamatoria.
Los extractos en n-hexano y cloroformo inhiben la producción de PGE2 y dicho
efecto se relaciona con la disminución en la expresión de COX-2.
El fraccionamiento del extracto en cloroformo mostró que la mayoría de sus
compuestos son de carácter polar y la viabilidad celular de los macrófagos
murinos frente a las fracciones es alta, excepto las fracciones Tc-F4 y Tc-F8
que fueron tóxicas para los macrófagos murinos.

GRUPO POLIFENOLES
59
Los resultados de la marcha fitoquímica mostraron que hay posibles
metabolitos secundarios, tales como glicósidos cardiotónicos, lactonas
sesquiterpenicas y quinonas. Las quinonas se han identificado en otras
especies de la familia Bignoniaceae.
La fracción F13 y F16 del extracto en cloroformo presentan la mayor actividad
anti-inflamatoria.
El fraccionamiento de la fracción F16 mostró que la mayoría de sus
compuestos son de carácter polar.
El análisis por medio de espectroscopia de resonancia magnética nuclear de
1H y 13C mostró que la fracción F16-3-b podría contener una mezcla de
derivados de ácido benzoico.

GRUPO POLIFENOLES
60
6 RECOMENDACIONES
Evaluar la actividad antioxidante y antibacteriana de los extractos de
T.chrysantha, teniendo en cuenta los resultados obtenidos en la presente
investigación y los reportes de estas actividades en la literatura para otras
especies de Tabebuia.
Evaluar la actividad anti-inflamatoria de la fracción F16-3-b.
Realizar nuevamente el fraccionamiento del extracto en cloroformo con una
mayor cantidad de masa.
Purificar las fracciones obtenidas del fraccionamiento del extracto en
cloroformo con una mezcla de solventes de menor polaridad.

GRUPO POLIFENOLES
61
7 BIBLIOGRAFIA
1. Daniell HW. Gastroesophageal reflux disease stimulation of nsaid-associated atrial fibrillation. Archives of Internal Medicine. 2011;171(7):702-10.
2. Gómez HA, González KN, Medina JD. Actividad Antiinflamatoria de Productos Naturales2011.
3. Molina SC. Farmacología en enfermería: Elsevier; 2007.
4. Ichiyanagu C. Epidemiología de la hemorragia digestiva. Acta Médica Peruana. 2006;23:152-5.
5. Lourenço JA, Pitangui CP, Jordão AA, Vannucchi H, Cecchi AO. Ausência de mutagenicidade e antimutagenicidade do extrato obtido das flores do ipê roxo [Tabebuia impetiginosa (Mart. ex DC.) Standl.]. Revista Brasileira de Plantas Medicinais. 2010;12:414-20.
6. Sathiya M, Muthuchelian K. Studies on Phytochemical Profile and Antibacterial Activity of Ethanolic Leaf Extract of Tabebuia rosea (Bertol.) DC. Ethnobotanical Leaflets. 2008;12:1153-7.
7. Gupta MP, Farmacéutica CSdQF, Bello SEPdCA. 270 plantas medicinales iberoamericanas: Programa Iberoamericano de Ciencia y Tecnología para el Desarrollo, Subprograma de Química Fina Farmacéutica; 1995.
8. Nakano K, Maruyama K, Murakami K, Takaishi Y, Tomimatsu T. Iridoids from Tabebuia avellanedae. Phytochemistry. 1993;32(2):371-3.
9. Böhler T, Nolting J, Gurragchaa P, Lupescu A, Neumayer H-H, Budde K, et al. Tabebuia avellanedae extracts inhibit IL-2-independent T-lymphocyte activation and proliferation. Transplant Immunology. 2008;18(4):319-23.
10. Byeon SE, Chung JY, Lee YG, Kim BH, Kim KH, Cho JY. In vitro and in vivo anti-inflammatory effects of taheebo, a water extract from the inner bark of Tabebuia avellanedae. Journal of Ethnopharmacology. 2008;119(1):145-52.
11. Kim BH, Lee J, Kim KH, Cho JY. Regulation of macrophage and monocyte immune responses by water extract from the inner bark of Tabebuia avellanedae. Journal of Medicinal Plants Research 2010;4(6):431-8.
12. Twardowschy A, Freitas CS, Baggio CH, Mayer B, dos Santos AC, Pizzolatti MG, et al. Antiulcerogenic activity of bark extract of Tabebuia avellanedae, Lorentz ex Griseb. Journal of Ethnopharmacology. 2008;118(3):455-9. Epub 2008/06/27.
13. Pereira E, Machado T, Leal I, Jesus D, Damaso C, Pinto A, et al. Tabebuia avellanedae naphthoquinones: activity against methicillin-resistant staphylococcal strains, cytotoxic activity and in vivo dermal irritability analysis. Annals of Clinical Microbiology and Antimicrobials. 2006;5(1):5.

GRUPO POLIFENOLES
62
14. Park B-S, Lee K-G, Shibamoto T, Lee S-E, Takeoka GR. Antioxidant Activity and Characterization of Volatile Constituents of Taheebo (Tabebuia impetiginosa Martius ex DC). Journal of Agricultural and Food Chemistry. 2003;51(1):295-300.
15. de Santana CF, de Lima O, d' Albuquerque IL, Lacerda AL, Martins DG. [Antitumoral and toxicological properties of extracts of bark and various wood components of Pau d'arco (Tabebuia avellanedae)]. Rev Inst Antibiot (Recife). 1968;8(1):89-94. Epub 1968/12/01. Observacoes sobre as propriedades antitumorais e toxicologicas do extra to do liber e de alguns componentes do cerne do Pau d'arco (Tabebuia avellanedae.
16. Méndez JM. Manejo de semillas de 100 especies forestales de América Latina: CATIE; 2000.
17. Ali RM, Houghton PJ, Hoo TS. Antifungal activity of some Bignoniaceae found in Malaysia. Phytotherapy Research. 1998;12(5):331-4.
18. Consoli RAGB, Mendes NM, Pereira JP, Santos BdS, Lamounier MA. Influência de diversos derivados de vegetais na sobrevida das larvas de Aedes fluviatilis(Lutz) (Diptera: Culicidade) em laboratorio. Memórias do Instituto Oswaldo Cruz. 1988;83:87-93.
19. Justiniano MJ, Fredericksen TS, Nash D. Ecología y silvicultura de especies menos conocidas. Tajibos o lapachos.: Proyecto de Manejo Forestal Sostenible (BOLFOR); 2000.
20. Almeida ECd, Menezes H. Anti-inflammatory activity of propolis extracts: a review. Journal of Venomous Animals and Toxins. 2002;8:191-212.
21. Calixto JB, Otuki MF, Santos ARS. Anti-Inflammatory Compounds of Plant Origin. Part I. Action on Arachidonic Acid Pathway, Nitric Oxide and Nuclear Factor κ B (NF-κB). Planta Med. 2003;69(11):973-83.
22. Kumar V, Cotran RS, Robbins SL. Patología humana: Elsevier Health Sciences; 2008.
23. Seibert K, Zhang Y, Leahy K, Hauser S, Masferrer J, Perkins W, et al. Pharmacological and biochemical demonstration of the role of cyclooxygenase 2 in inflammation and pain. Proceedings of the National Academy of Sciences of the United States of America. 1994;91(25):12013-7. Epub 1994/12/06.
24. Beca T, Hernández G, Bascones A. AINEs como tratamiento coadyuvante de la enfermedad periodontal. Avances en Periodoncia e Implantología Oral. 2007;19:101-13.
25. Kim YK, Kim RG, Park SJ, Ha JH, Choi JW, Park HJ, et al. In Vitro Antiinflammatory Activity of Kalopanaxsaponin A Isolated from Kalopanax pictus in Murine Macrophage RAW 264.7 Cells. Biological and Pharmaceutical Bulletin. 2002;25(4):472-6.

GRUPO POLIFENOLES
63
26. Villena AA, Regueiro JR, Larrea CL. Inmunología: Editorial Complutense; 1995.
27. Rang HP. Rang y Dale, farmacología: Elsevier; 2008.
28. López JL, Gulias A, Gutiérrez C, Cárdenas MdC, Carrillo E, Terrazas H, et al. Actitud ante los inhibidores selectivos de COX-2. Revista de investigación clínica. 2005;57:6-12.
29. Raghav SK, Gupta B, Agrawal C, Goswami K, Das HR. Anti-inflammatory effect of Ruta graveolens L. in murine macrophage cells. Journal of Ethnopharmacology. 2006;104(1–2):234-9.
30. Victoria MdC, Morón F, Morejón Z, Martínez MJ, López M. Tamizaje fitoquímico, actividad antiinflamatoria y toxicidad aguda de extractos de hojas de Annona squamosa L. Revista Cubana de Plantas Medicinales. 2006;11:0-0.
31. Olmstead RG, Zjhra ML, Lohmann LG, Grose SO, Eckert AJ. A molecular phylogeny and classification of Bignoniaceae. American journal of botany. 2009;96(9):1731-43.
32. Blair S. Plantas antimaláricas de Tumaco: Costa Pacífica colombiana: Universidad de Antioquia; 2005.
33. Flora de la Real Expedición Botánica del Nuevo Reino de Granada: Ed. Cultura Hispánica; 1985.
34. Girard M, Kindack D, Dawson BA, Ethier J-C, Awang DVC, Gentry AH. Naphthoquinone Constituents of Tabebuia spp. Journal of Natural Products. 1988;51(5):1023-4.
35. Rao MM, Kingston DGI. Plant Anticancer Agents. XII. Isolation and Structure Elucidation of New Cytotoxic Quinones From Tabebuia cassinoides. Journal of Natural Products. 1982;45(5):600-4.
36. Linardi MdCF, De Oliveira MM, Sampaio MRP. Lapachol derivative active against mouse lymphocytic leukemia P-388. Journal of Medicinal Chemistry. 1975;18(11):1159-61.
37. Koyama J, Morita I, Tagahara K, Hirai K-I. Cyclopentene dialdehydes from Tabebuia impetiginosa. Phytochemistry. 2000;53(8):869-72.
38. Otero R, Núñez V, Jiménez SL, Fonnegra R, Osorio RG, García ME, et al. Snakebites and ethnobotany in the northwest region of Colombia: Part II: Neutralization of lethal and enzymatic effects of Bothrops atrox venom. Journal of Ethnopharmacology. 2000;71(3):505-11.
39. Camacho RL. Manual de identificación de especies no maderables del corregimiento de Tarapacá Colombia: Instituto Amazónico de Investigaciones Científicas; 2006.

GRUPO POLIFENOLES
64
40. Bueno J, Isaza G, Gutiérrez F, Carmona W, Pérez J. Estudio etnofarmacológico de plantas usadas empíricamente por posibles efectos inmunoestimulantes. . Rev Med de Risaralda 2001;7(1):7-11.
41. Isaza G, Bueno J, Jaramillo R, Gutiérrez F, Guzmán A. Estudio etnofarmacológico de plantas usadas empíricamente en 4 ciudades del centro-occidente colombiano. Medomai. 2001;2(1):8-9.
42. Perez JE, Isaza G, Bueno JG, Arango MC, Hincapié BL, Nieto AM, Londoño DP. Efecto de los extractos de Phenax rugosus, Tabebuia chrysantha, Althernantera illiamsii y Solanum dolichosepalum sobre el leucograma y la producción de anticuerpos en ratas. Revista medica de Risaralda. 2004;10 (2):13-21.
43. Ospina LF, Aragon DM, Vergel NE, Isaza G, Pérez JE. Anti-inflammatory and antioxidant activities of Phenax rugosus (Poir.) Wedd and Tabebuia chrysanta G. Nicholson. Vitae. 2011;18:49-55.
44. Zani CL, De Oliveira AB, De Oliviera GG. Furanonaphthoquinones from Tabebuia ochracea. Phytochemistry. 1991;30(7):2379-81.
45. B. de Oliveira A, Raslan DS, G. de Oliveira G, Maia JGS. Lignans and naphthoquinones from Tabebuia incana. Phytochemistry. 1993;34(5):1409-12.
46. Díaz F, Medina JD. Furanonaphthoquinones from Tabebuia ochracea ssp. neochrysanta. Journal of Natural Products. 1996;59(4):423-4.
47. Sharma PK, Khanna RN, Rohatgi BK, Thomson RH. Tecomaquinone-III: A new quinone from Tabebuia pentaphylla. Phytochemistry. 1988;27(2):632-3.
48. Manners GD, Jurd L. A new naphthaquinone from Tabebuia guayacan. Phytochemistry. 1976;15(1):225-6.
49. Wagner H, Kreher B, Lotter H, Hamburger MO, Cordell GA. Structure Determination of New Isomeric Naphtho[2,3-b]furan-4,9-diones from Tabebuia avellanedae by the selective-INEPT technique. Helvetica Chimica Acta. 1989;72(4):659-67.
50. Blatt CTT, Salatino A, Salatino MLF. Flavonoids of Tabebuia caraiba (Bignoniaceae). Biochemical Systematics and Ecology. 1996;24(1):89.
51. Warashina T, Nagatani Y, Noro T. Constituents from the Bark of Tabebuia impetiginosa. Chemical and Pharmaceutical Bulletin. 2006;54(1):14-20.
52. de Almeida ER, da Silva Filho AA, Dos Santos ER, Correia Lopes CA. Antiinflammatory action of lapachol. Journal of Ethnopharmacology. 1990;29(2):239-41.
53. Subramanian S, Ferreira MMC, Trsic M. A Structure-Activity Relationship Study of Lapachol and Some Derivatives of 1,4-Naphthoquinones Against Carcinosarcoma Walker 256. Struct Chem. 1998;9(1):47-57.

GRUPO POLIFENOLES
65
54. Dolan ME, Frydman B, Thompson CB, Diamond AM, Garbiras BJ, Safa AR, et al. Effects of 1,2-naphthoquinones on human tumor cell growth and lack of cross-resistance with other anticancer agents. Anti-cancer drugs. 1998;9(5):437-48.
55. Renou SG, Asis SE, Abasolo MI, Bekerman DG, Bruno AM. Monoarylhydrazones of alpha-lapachone: synthesis, chemical properties and antineoplastic activity. Die Pharmazie. 2003;58(10):690-5.
56. Byung C, Jun L, KO Woo S, Young K, Yung C, Ho-Sung K, et al. Anti-inflammatory effects of aqueous extract from Dichroa febrifuga root in rat liver. Acta Pharcologica sinica. 2003;24 (2):127-32.
57. Burnett AR, Thomson RH. Naturally occurring quinones. Part XII. Extractives from Tabebuia chrysantha nichols and other bignoniaceae. Journal of the Chemical Society C: Organic. 1968(0):850-3.
58. Awang DVC, Dawson BA, Ethier J-C, Gentry AH, Girard M, Kindack D. Naphthoquinone Constituents of Commercial Lapacho/Pau d'arco/Taheebo Products. Journal of Herbs, Spices & Medicinal Plants. 1995;2(4):27-43.
59. Roitt I BJ, Male D, Roth D. Inmunología: Elsevier España; 2007.
60. Chen K, Pittman RN, Popel AS. Nitric oxide in the vasculature: where does it come from and where does it go? A quantitative perspective. Antioxidants & redox signaling. 2008;10(7):1185-98.
61. Reumatología SEd. Artrosis: Fisiopatología, diagnóstico y tratamiento: Editorial Medica Panamericana Sa de; 2010.
62. Tamargo J, Caballero R, Gómez R, Núñez L, Vaquero M, Delpón E. Efectos del óxido nítrico sobre la función cardíaca. Revista Española de Cardiología. 2006;6(Supl.A):3-20.
63. Anderson GL, McNellis LA. Enzyme-Linked Antibodies: A Laboratory Introduction to the ELISA Assay. Journal of Chemical Education. 1998;75(10):1275.
64. Losada FL. Fundamentos moleculares en medicina: El Manual Moderno; 2005.
65. Tintinalli JE, Stapczynski JS. Tintinalli's Emergency Medicine: A Comprehensive Study Guide: McGraw-Hill; 2011.
66. Bilbao MdR. Análisis fitoquímico preliminar: Oficina de publicaciones Universidad del Quindío; 1997.
67. Dominguez XA. Métodos de investigación fitoquímica: Editorial Limusa; 1973.
68. INEP CP RAS NO, Commerce CCcbtUSSo, reserved. obotUSoAAr. p-Benzoquinone. 2007.

GRUPO POLIFENOLES
66
69. Concepción J, Puerta B, P C, Ureña P. Prácticas de biología molecular: Pontificia Universidad Javeriana.
70. Escrig AJ. Manual de Neurogenética: Díaz de Santos; 2003.
71. Yoon WJ, Ham YM, Yoo B-S, Moon JY, Koh J, Hyun CG. Oenothera laciniata inhibits lipopolysaccharide induced production of nitric oxide, prostaglandin E2, and proinflammatory cytokines in RAW264.7 macrophages. Journal of Bioscience and Bioengineering. 2009;107(4):429-38.
72. Hay C, de Belleroche J. Carrageenan-induced hyperalgesia is associated with increased cyclo-oxygenase-2 expression in spinal cord. NeuroReport. 1997;8(5):1249-51.
73. Garcez FR, Garcez WS, Mahmoud TS, Figueiredo PdO, Resende UM. Novos constituintes químicos das cascas do caule de Tabebuia heptaphylla. Química Nova. 2007;30:1887-91.

GRUPO POLIFENOLES
67
ANEXO 1
Primers Sequence
Sense mCOX2 5’-GGA GAG ACT ATC AAG ATA GTG ATC-3’
Antisense mCOX2 5’-ATG GTC AGT AGA CTT TTA CAG CTA-3’
Sense mbactin 5’-TGT GAT GGT GGG AAT GGG TCA G-3’
Antisense mbactin 5’-TTT GAT GTC ACG CAG GAT TTC C-3’
Forward miNOS 5’-CCC TTC CGA AGT TTC TGG CAG CAG C-3’
Reverse miNOS 5’-GGC TGT CAG AGC CTC GTG GCT TTG G-3’
Forward mTNF-α 5’-TTG ACC TCA GCG CTG AGT TG-3’
Reverse mTNF-α 5’-CCT GTA GCC CAC GTC GTA GC-3’

GRUPO POLIFENOLES
68
ANEXO 2
Cromatograma del extracto en n-hexano
Espectro UV/VIS del pico 1

GRUPO POLIFENOLES
69
Espectro UV/VIS del pico 7

GRUPO POLIFENOLES
70
ANEXO 3
Cromatograma del extracto en butanol
Espectro UV/VIS del pico 2
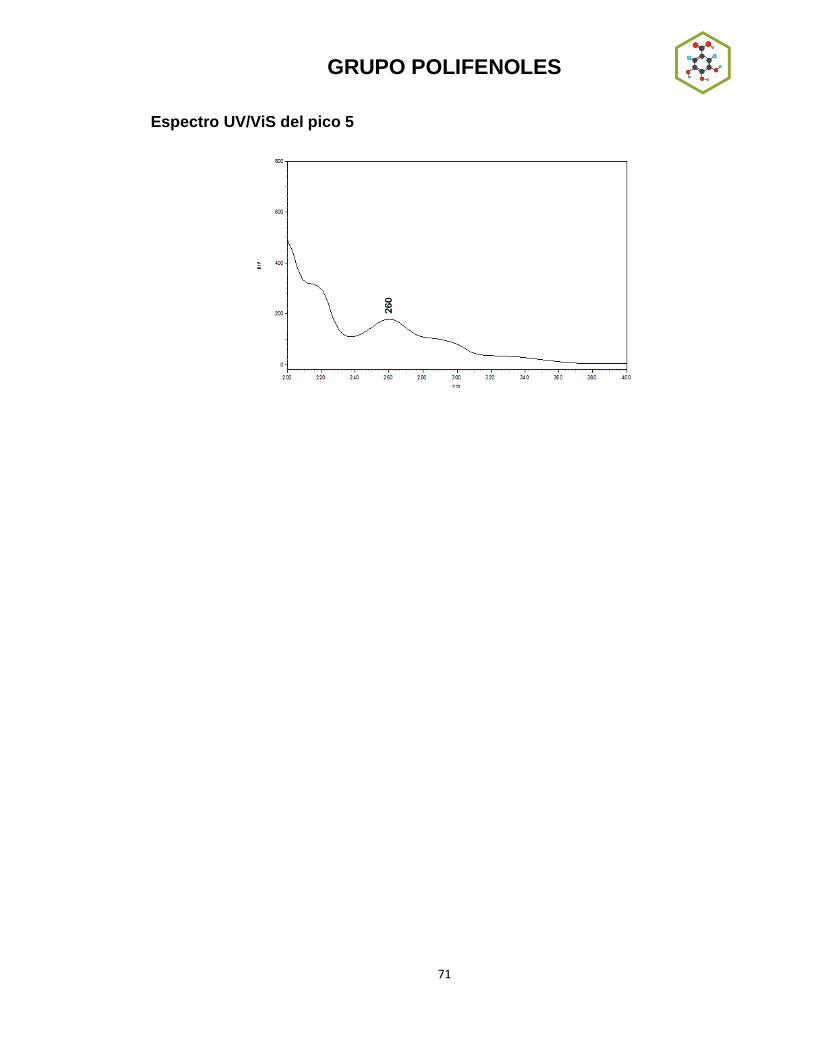
GRUPO POLIFENOLES
71
Espectro UV/ViS del pico 5

GRUPO POLIFENOLES
72
ANEXO 4
Cromatograma del extracto en agua
Espectro UV/VIS del pico 1

GRUPO POLIFENOLES
73
Espectro UV/VIS del pico 2

GRUPO POLIFENOLES
74
ANEXO 5
Viabilidad celular de los macrófagos RAW264.7 frente a los extractos de
corteza interna de T. chrysantha.
Extractos
Muestra Concentración (μg/mL) Viabilidad (%)
n-Hexano
2,5 79
1,2 78
0,6 130
Cloroformo
2,3 113
1,2 104
0,6 139
Acetato de etilo
2,4 98
1,2 98
0,6 129
Butanol
2,1 64
1,1 89
0,5 115
Agua
2,2 103
1,1 82
0,6 92
Controles
Control Viabilidad (%)
Control positivo con células 100
Control negativo sin células 0

GRUPO POLIFENOLES
75
ANEXO 6
Efecto inhibitorio de los extractos de la corteza interna de T. chrysantha
sobre la producción de PGE2 en macrófagos murinos estimulados con LPS.
Muestra Inhibición (%)
n-Hexano 23,5
Cloroformo 47,1
Acetato de etilo 30,8
Butanol 5,6
Agua 34,3
LPS (-) 100,0
LPS (+) 0,0
Asp, LPS (+) 2,5
Asp, DUP, LPS (+) 88,3

GRUPO POLIFENOLES
76
ANEXO 7
Efecto inhibitorio de los extractos de la corteza interna de T. chrysantha
sobre la producción de TNF-α en macrófagos murinos estimulados con LPS
Muestra Inhibición (%)
n-Hexano 112,8
Cloroformo 111,8
Acetato de etilo 121,7
Butanol 71,0
Agua 101,4
LPS (-) 100,0
LPS (+) 0,0

GRUPO POLIFENOLES
77
ANEXO 8
Efecto inhibitorio de los extractos de la corteza interna de T. chrysantha
sobre la producción de NO en macrófagos murinos estimulados con LPS
Muestra Inhibición (%)
n-Hexano -312,2
Cloroformo -13,3
Acetato de etilo -139,0
Butanol 59,6
Agua -187,7
LPS (-) 100,0
LPS (+) 0,0
GRUPO POLIFENOLES
78
ANEXO 9
Efecto inhibitorio de las expresiones de mRNA de COX-2, TNF-α e iNOS
sobre macrófagos murinos RAW264.7 estimulados con LPS
Muestra COX-2
(Rf)
TNFα
(Rf)
iNOS
(Rf)
n-Hexano 0,04 1,93 0,99
Cloroformo 0,07 1,33 0,70
Acetato de etilo 0,17 0,95 0,56
Butanol 0,15 1,26 0,70
Agua 0,18 1,91 1,05
LPS (-) 0,06 1,22 0,64
LPS (+) 0,14 1,15 0,65
Asp, LPS (+) 0,04 1,66 0,85
Asp, DUP, LPS (+) 0,14 1,33 0,74