abstract - iai.asm.orgiai.asm.org/content/early/2011/06/27/iai.05226-11.full.pdf4 enrique llobet,1,2...
TRANSCRIPT

- 1 -
Analysis of the networks controlling the antimicrobial peptide-dependent induction of 1
Klebsiella pneumoniae virulence factors. 2
3
Enrique Llobet,1,2 Miguel A. Campos1, Paloma Giménez1,2, David Moranta1,2; José A. 4
Bengoechea1,2,3,* 5
6
Laboratory Microbial Pathogenesis, Fundació d'Investigació Sanitària de les Illes Balears (FISIB), 7
Recinto Hospital Joan March, 07110, Bunyola, Spain1 8
Program Host-Pathogen interactions, Centro de Investigación Biomédica en Red Enfermedades 9
Respiratorias (CibeRes), Bunyola, Spain2 10
Consejo Superior de investigaciones Científicas (CSIC), Madrid, Spain3 11
12
*Corresponding author: 13
Laboratory Microbial Pathogenesis, 14
Fundació d'Investigació Sanitària de les Illes Balears (FISIB) 15
Recinto Hospital Joan March 16
Carretera Soller Km 12 17
07110 Bunyola 18
Spain 19
Phone: +34 971 011780 Fax: +34 971 011797 20
E-mail: [email protected] 21
22
Keywords: Klebsiella, antimicrobial peptides, capsule polysaccharide, lipid A 23
Running title: Polymyxin induces Klebsiella pneumoniae countermeasures 24
25
26
27
Copyright © 2011, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.Infect. Immun. doi:10.1128/IAI.05226-11 IAI Accepts, published online ahead of print on 27 June 2011
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 2 -
ABSTRACT 28
Antimicrobial peptides (APs) impose a threat to the survival of pathogens and it is reasonable to 29
postulate that bacteria have developed strategies to counteract them. Polymyxins are becoming the 30
last resort to treat infections caused by multi-drug resistant Gram-negative bacteria and, similar to 31
APs, they interact with the anionic lipopolysaccharide. Given that polymyxins and APs share the 32
initial target, it is possible that bacterial defense mechanisms against polymyxins will be also 33
effective against host APs. We sought to determine whether exposure to polymyxin will increase 34
Klebsiella pneumoniae resistance to host APs. Indeed, exposure of K. pneumoniae to polymyxin 35
induces cross-resistance to polymyxin itself but also to APs present in the airways. Polymyxin 36
treatment up-regulates the expression of the capsule polysaccharide operon and the loci required to 37
modify the lipid A with aminoarabinose and palmitate with a concomitant increase in capsule and 38
lipid A species containing such modifications. Moreover, these surface changes contribute to APs 39
resistance and also to polymyxin-induced cross resistance to APs. Bacterial loads of lipid A mutants 40
in trachea and lungs of intranasally infected mice were lower than those of wild-type strain. PhoPQ, 41
PmrAB and Rcs system govern polymyxin-induced transcriptional changes and there is a cross-talk 42
between PhoPQ and Rcs system. Our findings support the notion that Klebsiella activates a defense 43
program against APs which is controlled by three signaling systems. Therapeutic strategies directed 44
to prevent the activation of this program could be a new approach worthy exploring to facilitate the 45
clearance of the pathogen from the airways. 46
47
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 3 -
INTRODUCTION 48
Antimicrobial peptides (APs) are ubiquitous in nature and in vertebrates they belong to the 49
arsenal of weapons of the innate immune system against infections. There are four structural classes 50
of APs: the disulfide-bonded β-sheet peptides, the amphipathic α-helical peptides, the extended 51
peptides and the loop-structured peptides (8,32,50). Despite their diverse size and structures, nearly 52
all APs have a net positive charge and the three dimensional folding results in an amphipathic 53
structure (8,32,50). In most cases, the action of APs is initiated through electrostatic interaction with 54
the bacterial surface (8,32,50,62) and, in the case of Gram-negative bacteria, APs interact with the 55
anionic lipid A moiety of the lipopolysaccharide (LPS) (8,32,50,62). 56
APs impose a threat to the survival of pathogens and therefore it is reasonable to postulate 57
that bacteria have developed means to sense the presence of APs in order to activate 58
countermeasures to limit their effectiveness. Furthermore, given the importance of APs in host 59
defense, it is likely that these countermeasures will be important virulence factors. Bacteria utilize 60
phosphorelay signaling cascades in the form of two-component systems to respond and adapt to 61
different hostile environments. The sensors of these two component systems respond to particular 62
cues by modulating the phosphorylation status of their cognate regulators which are often 63
transcription factors. As a result, genes necessary for growth and survival are up-regulated whereas 64
genes deleterious for infectivity might be down-regulated. It can be speculated that bacteria may 65
utilize two-component systems to transduce AP-mediated signals hence leading to the activation of 66
bacterial defense mechanisms. Supporting this idea, Salmonella PhoPQ two-component system 67
regulates genes necessary for intracellular survival, cellular invasion, and it is required for 68
resistance to a subset of APs (6,20,21,25,26). 69
Polymyxins B (PxB) and E (colistin) are two antibiotics originally derived from Bacillus 70
polymyxa and made available for clinical use in the late 1950s and early 1960s. Polymyxins are 71
pentacationic amphipathic lipopeptide antibiotics characterized by a heptapeptide ring and a fatty 72
acid tail (63). Polymyxins are active against Gram-negative bacteria and, similar to APs, they do 73
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 4 -
interact with the anionic LPS. Soon after their introduction, the clinical use was limited due to 74
perceived toxic side effects and the emergence of new antimicrobials (17,35). However, the 75
occurrence of multidrug-resistant Gram-negative bacteria has prompted to reconsider polymyxin 76
therapies (24,63). Nevertheless, the pharmacokinetics and pharmacodynamics of polymyxins are 77
poorly understood making it possible that bacteria are exposed to sublethal concentrations during 78
treatment. Consequently, the possibility exists that bacteria may activate defense mechanisms 79
against polymyxins. Furthermore, given that polymyxins and APs share the initial target, it is 80
possible that PxB countermeasures will be also effective against host APs thus contributing to 81
bacterial resistance and survival in host tissues. 82
To study this hypothesis, in this work we used the Gram-negative human pathogen K. 83
pneumoniae. The frequent isolation of K. pneumoniae multidrug resistant strains makes polymyxins 84
a therapeutic option (24). There is paucity of information on the mechanisms of resistance of this 85
pathogen against polymyxins and APs. By mass spectrometry and genetic methods, we demonstrate 86
that PxB indeed induces the expression of loci conferring resistance against PxB but also against 87
host APs. We demonstrate that these loci play an important role in K. pneumoniae virulence. 88
Finally, we show that at least three signaling transduction systems govern PxB-induced changes. 89
90
MATERIAL AND METHODS 91
Bacterial strains and growth conditions. 92 93 Bacterial strains and plasmids used in this study are listed in Table 1. Strains were grown in Luria–94
Bertani (LB) at 37ºC. When appropriate, antibiotics were added to the growth medium at the 95
following concentrations: rifampicin (Rif) 25 µg/ml, ampicillin (Amp), 100 µg/ml for K. 96
pneumoniae and 50 µg/ml for E. coli; kanamycin (Km) 100 µg/ml; chloramphenicol (Cm) 12.5 97
µg/ml. 98
K. pneumoniae 52145 mutant construction. 99
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 5 -
Primers for mutant construction (Table 2) were designed based on the available genome sequence 100
of K. pneumoniae subsp. pneumoniae MGH78578 (GenBank CP000647.1). DNA fragments for 101
phoPQ, rcsB, and pagP were PCR-amplified, gel-purified and cloned into pGEM-T Easy 102
(Promega) to obtain pGEMTphoPQ, pGEMTrcsB, and pGEMTpagP respectively. These plasmids 103
were amplified by inverse PCR using the method described by Byrappa et al (9) to delete internal 104
coding regions of phoQ, rcsB, and pagP. A kanamycin resistance cassette, obtained as a 1.4 kb PstI 105
blunt-ended fragment from pUC-4K (Pharmacia), was cloned into the plasmids obtained by inverse 106
PCR to generate pGEMTΔphoPQGB and pGEMTΔpagPGB. ΔphoPQ::GB, ΔrcsB, and ΔpagP::GB 107
alleles were obtained by PvuII digestion of pGEMTΔphoPQGB, pGEMTΔrcsB, and 108
pGEMTΔpagPGB respectively, gel-purified and cloned into SmaI-digested pMAKSACB. 109
pMAKSACB is a suicide vector that carries a rep101ts origin of replication, an oriT sequence for 110
conjugational transfer and a Cm resistance marker (19). It also carries the sacB gene that mediates 111
sucrose sensitivity as a positive selection for the excision of the vector after double crossing-over 112
(19). pMAKSACΔphoPQGB, pMAKSACΔrcsB, and pMAKSACΔpagPGB, were electroporated 113
into E. coli S17-1λpir, from which the plasmids were mobilized into K. pneumoniae 52145. 114
Transconjugates were selected after growth on LB plates supplemented with Cm at 30oC. Bacteria 115
from 10 individual colonies were pooled in 500 μl PBS, serially diluted in PBS, and spread on LB 116
plates with Cm which were incubated at 42ºC in order to select merodiploids in which the suicide 117
vector was integrated into the chromosome by homologues recombination. 5-10 merodiploids were 118
serially diluted in PBS and dilutions spread in LB plates without NaCl containing 10% sucrose 119
which were incubated at 30oC. The recombinants that survived 10% sucrose were checked for their 120
antibiotic resistance and the appropriate replacement of the wild-type alleles by the mutant ones was 121
confirmed by PCR (data not shown). Recombinants selected were named 52145-∆phoQGB; 52145-122
∆rcsB, and 52145-∆pagPGB. To confirm that pagP mutation has no polar effects, the expression of 123
the downstream gene, cpsE, was analyzed by real time (RT) quantitative PCR (RT-qPCR). Briefly, 124
200 ng cDNA, obtained by retrotranscription of 2 μg of total RNA using a commercial RT2 First 125
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 6 -
Strand kit (Superarray Bioscience Corporation), were used as a template in a 25-μl reaction mixture 126
containing 1x SYBR green RT2 qPCR Master Mix (Superarray Bioscience Corporation) and primer 127
mix. rpoD was amplified as control. RT-qPCR analyses were performed as previously described 128
(48). The expression of cpsE by 52145-∆pagPGB was similar to that by the wild-type strain (data 129
not shown). 130
DNA fragments for pmrAB, and pmrF were PCR-amplified, gel-purified and cloned into pGEM-T 131
Easy (Promega) to obtain pGEMTpmrAB, and pGEMTpmrF respectively. These plasmids were 132
amplified by inverse PCR to delete internal coding regions of pmrAB, and pmrF respectively. A 133
kanamycin resistance cassette, obtained as a 1.5 kb PCR fragment from pKD4 (14), was cloned into 134
the plasmids obtained by inverse PCR to generate pGEMTΔpmrABKm, and pGEMTΔpmrFKm. 135
ΔpmrAB::Km, and ΔpmrF::Km alleles were obtained by PvuII digestion of pGEMTΔpmrABKm, 136
and pGEMTΔpmrFKm respectively, and cloned into SmaI-digested pKOV (42). Recombinants in 137
which the wild-type allele was replaced by the mutant one were selected as described previously 138
and named 52145-∆pmrABKm, and 52145-∆pmrFKm. The kanamycin cassette was excised by Flp-139
mediated recombination using plasmid pFLP2 (37) and the generated mutants were named 52145-140
∆pmrAB, and 52145-∆pmrF. RT-qPCR analysis revealed that the expression of pmrI, pmrF 141
downstream gene, was not significantly different between pmrF mutant and the wild-type strain 142
(data not shown). 143
To obtain a pmrD mutant, two sets of primers (Table 2) were used to obtain two different pmrD 144
fragments, PmrDUP, and PmrDDown. These fragments were annealed at their overlapping region 145
and amplified by PCR as a single fragment which was cloned into pGEM-T Easy to obtain 146
pGEMT∆pmrD. A kanamycin cassette was PCR amplified from pKD4 and cloned into pGEM-T 147
Easy to give pGEMTFRTKM. The cassette was obtained as a BamHI fragment which was cloned 148
into BamHI-digested pGEMT∆pmrD to generate pGEMTΔpmrDKm. ΔpmrD::Km allele was gel-149
purified after PvuII digestion of pGEMTΔpmrDKm, and cloned into SmaI-digested pKOV (42). 150
Recombinants in which the wild-type allele was replaced by the mutant one were selected as 151
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 7 -
described previously and named 52145-∆pmrDkm. The kanamycin cassette was excised by Flp-152
mediated recombination using plasmid pFLP2 (37) and the generated mutant was named 52145-153
∆pmrD. 154
52145-∆phoQGB-∆pmrAB, and 52145-∆phoQGB-∆rcsB double mutants were obtained mobilizing 155
the pMAKSACΔphoPQGB plasmid into 52145-∆pmrAB, and 52145-∆rcsB, respectively. The 156
replacement of the wild-type alleles by the mutant ones was done as described above and confirmed 157
by PCR (data not shown). 158
159
Construction of reporter fusions. 160
DNA fragments containing the promoter regions of the cps, pmrH, pagP, mgtA, pmrD, phoP, ugd, 161
rcsD and rcsC genes were amplified by PCR using Vent polymerase, EcoRI-digested, gel-purified 162
and cloned into EcoRI-SmaI digested pGPL01 suicide vector (29). This vector contains a 163
promoterless firefly luciferase gene (lucFF) and a R6K origin of replication. Plasmids in which 164
lucFF was under the control of the Klebsiella promoters were identified by restriction digestion 165
analysis and named pGPLKpnPcps, pGPLKpnPmrH, pGPLKpnPagP, pGPLKpnPmgtA, 166
pGPLKpnPmrD, pGPLKpnPphoP, pGPLKpnPugd, pGPLKpnPrcsD, and pGPLKpnPrcsC, 167
respectively. Plasmids were electroporated into the different Klebsiella strains used in this study. 168
Strains in which the suicide vector was integrated into the genome by homologous recombination 169
were selected. This was confirmed by Southern blot (data not shown). 170
171
Luciferase activity. 172
The reporter strains were grown on an orbital incubator shaker (180 r.p.m.) until late log phase and, 173
when required, PxB (1 μg/ml) was added and the culture incubated for 1 h more. At the end of the 174
incubation the OD at 540 nm was recorded. A 100 µl aliquot of the bacterial suspension was mixed 175
with 100 µl of luciferase assay reagent (1 mM D-luciferin [Synchem] in 100 mM citrate buffer pH 176
5). Luminescence was immediately measured with a LB9507 Luminometer (Berthold) and 177
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 8 -
expressed as relative light units (RLU)/OD540. All measurements were carried out in quintuplicate 178
on at least three separate occasions. 179
180
Antimicrobial peptide susceptibility assay. 181
Bacteria were grown at 37ºC in 5 ml LB, harvested (2500 x g, 20 min, 24ºC) in the exponential 182
growth phase (OD600 0.6). When required, PxB (1 μg/ml) was added and the culture incubated for 1 183
h more. Bacteria were washed once with PBS and a suspension containing approximately 1x106 184
cfu/ml was prepared in 10 mM PBS (pH 6.5), 1 % Tryptone Soya Broth (TSB; Oxoid), and 100 185
mM NaCl. Aliquots (5 μl) of this suspension were mixed in 1.5 ml microcentrifuge tubes with 186
various concentrations of AP. In all cases the final volume was 30 μl. After 1 h incubation at 37oC, 187
the contents of the tubes were plated on LB agar. Colony counts were determined and results were 188
expressed as percentages of the colony count of bacteria not exposed to antibacterial agents. All 189
experiments were done with duplicate samples on at least four independent occasions. 190
The 50 % inhibitory concentration of AP (IC50) was defined as the concentration producing a 50 % 191
reduction in the colony counts compared with bacteria not exposed to the antibacterial agent. 192
Following guidelines of the National Institutes of Health Chemical Genomics Center 193
(www.ncgc.nih.gov), IC50 of a given AP was determined from dose-response curve data fit using a 194
standard four-parameter logistic nonlinear regression analysis. Dose-response experiments were 195
done on four independent occasions. Results are reported as mean ± SEM. 196
197
Isolation and analysis of lipid A. 198
Lipid As were extracted using an ammonium hydroxide/isobutyric acid method and subjected to 199
negative ion matrix-assisted laser desorption ionization time-of-flight (MALDI-TOF) mass 200
spectrometry analysis (16,53). Briefly, lyophilized bacteria (10 mg) were resuspended in 400 µl 201
isobutyric acid/1M ammonium hydroxide (5:3, v/v) and were incubated in a screw-cap test tube at 202
100ºC for 2 h, with occasional vortexing. Samples were cooled in ice water and centrifuged (2,000 203
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 9 -
x g for 15 min). The supernatant was transferred to a new tube, diluted with an equal volume of 204
water, and lyophilized. The sample was then washed twice with 400 µl methanol and centrifuged 205
(2,000 x g for 15 min). The insoluble lipid A was solubilized in 100-200 µl 206
chloroform/methanol/water (3:1.5:0.25, v/v/v). Analyses were performed on a Bruker Autoflex II 207
MALDI-TOF mass spectrometer (Bruker Daltonics, Incorporated) in negative reflective mode with 208
delayed extraction. The ion-accelerating voltage was set at 20 kV. To analyze the samples, few 209
microliters of lipid A suspension (1 mg/ml) were desalted with few grains of ion-exchange resin 210
(Dowex 50W-X8; H+) in a 1.5 ml microcentrifuge tube. 1 µl aliquot of the suspension (50–100 µl) 211
was deposited on the target and covered with the same amount of dihydroxybenzoic acid matrix 212
(Sigma Chemical Co., St. Louis, MO) dissolved in 0.1 M citric acid. Different ratios between the 213
samples and dihydroxybenzoic acid were used when necessary. Alternatively, lipid A was mixed 214
with 5-chloro-2-mercapto-benzothiazole (Sigma Chemical Co., St. Louis, MO) [20 mg/ml in 215
chloroform/methanol (1:1, v/v)] at a ratio of 1:5. Each spectrum was an average of 300 shots. A 216
peptide calibration standard (Bruker Daltonics) was used to calibrate the MALDI-TOF. Further 217
calibration for lipid A analysis was performed externally using lipid A extracted from E. coli strain 218
MG1655 grown in LB at 37oC. Interpretation of the negative-ion spectra is based on earlier studies 219
showing that ions with masses higher than 1000 gave signals proportional to the corresponding lipid 220
A species present in the preparation (3,41,52,58). Important theoretical masses for the interpretation 221
of peaks found in this study are: C14:OH, 226; C12, 182, C14, 210; aminoarabinose (Ara4N), 131; C16, 222
239. 223
224
Capsule polysaccharide (CPS) purification and quantification 225
Cell-associated CPSs from K. pneumoniae strains, grown in 5 ml LB, were obtained using the hot 226
phenol-water method exactly as previously described (10). CPS was quantified by determining the 227
concentration of uronic acid in the samples, using a modified carbazole assay (7), exactly as 228
described by Rahn and Whitfield (57). 229
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 10 -
Intranasal infection model 230
Six- to seven-week-old virus-free female C57BL/ 6JOlaHsd mice (Harlan) were anesthetized by 231
intraperitoneal injection with a mixture containing ketamine (50 mg/kg) and xylazine (5 mg/kg). 232
Bacteria were grown at 37ºC in 5 ml LB, harvested (2500 x g, 20 min, 24ºC) in the exponential 233
growth phase, resuspended in PBS, and adjusted to 1 x 106 cfu/ml. 20 μl of the bacterial suspension 234
were inoculated intranasally in four 5 μl aliquots. To facilitate consistent inoculations, mice were 235
held vertically during inoculation and placed on a 45° incline while recovering from anesthesia. At 236
the indicated times after infection, mice were euthanized by cervical dislocation. Trachea and lungs 237
were aseptically removed, weighed and homogenized in 500 μl PBS for bacterial load 238
determination. Results were reported as log CFU per gram of tissue. Mice were treated in 239
accordance with the European Convention for the Protection of Vertebrate Animals used for 240
Experimental and other Scientific Purposes (Directive 86/609/EEC) and in agreement with the 241
Bioethical Committee of the University of the Balearic Islands. 242
243
Statistical analysis. 244
Statistical analyses were performed using one-way analysis of variance (ANOVA) with Bonferroni 245
contrasts or the two-tailed t test or, when the requirements were not met, by the Mann-Whitney U 246
test. P < 0.05 was considered statistically significant. The analyses were performed using Prism4 for 247
PC (GraphPad Software). 248
249
250
251
252
253
254
255
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 11 -
RESULTS 256
Exposure of K. pneumoniae to polymyxin B increases the resistance to antimicrobial peptides. 257
We sought to determine whether exposure to PxB will increase the resistance of Kp52145, 258
wild-type strain, to PxB. Results shown in Figure 1A demonstrate that 1 h treatment with PxB (1 259
μg/ml) resulted in a markedly increased resistance to killing by this agent. To test whether exposure 260
to PxB also induces cross-resistance to other APs, killing assays were performed with human β-261
defensins (hBD) 1 and 2, HNP-1 and magainin II. hBD1 is constitutively expressed in the airways 262
(46). hBD2 is produced by airway epithelial cells upon induction by cytokines or by the presence of 263
pathogens (33,34,46,59) and its levels increase several folds in lung during pneumonia (36). HNP-1 264
is produced by neutrophils and released to the medium after degranulation (22). Magainin II is an 265
AP produced by frogs and it is widely used as a model AP (45). Bacteria treated with PxB were also 266
more resistant against hBD1, hBD2, HNP-1 and magainin II than non-treated bacteria (Fig 1B-E). 267
In summary, these observations demonstrate that K. penumoniae resistance to APs can be induced 268
by exposure to PxB. 269
270
Exposure of K. pneumoniae to polymyxin B increases capsule expression and LPS 271
modifications. 272
Recently, we have shown that K. pneumoniae CPS acts as a protective shield against APs 273
(10), whereas released CPS traps APs thereby blocking their bactericidal activity (43). Moreover, 274
there is a correlation between the amount of CPS and the resistance to PxB (10). Therefore, the 275
observed PxB-induced resistance could be mediated by an increase in CPS expression. Indeed, 276
exposure of Kp52145 to PxB (1 μg/ml) up-regulated the transcription of the cps operon (Fig 2A) 277
with a concomitant increased in the amount of cell-bound CPS (64.4 ± 3 μg/104 CFU versus 113 ± 278
5 μg/104 CFU, respectively; P < 0.05 [two-tailed t test]). To test whether PxB-induced resistance 279
was solely due to increase expression of CPS, we analyzed the effect of PxB on 52145-∆wcaK2, a 280
CPS mutant. Figure 2 demonstrates that exposure of 52145-∆wcaK2 to PxB also induced cross-281
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 12 -
resistance to PxB (Fig 2B), hBD1 (Fig 2C), hBD2 (Fig 2D), HNP-1 (Fig 2E) and magainin II (Fig 282
2F). To compare the PxB-induced levels of resistance against the different APs between Kp52145 283
and 52145-∆wcaK2, we determined the IC50 of APs for these strains pre-treated with PxB. IC50 of 284
PxB, hBD1, hBD2, HNP-1, and magainin II for PxB-treated Kp52145 were 4.5±0.8, 19±1.1, 285
10.2±0.6, 48±2.1 and 56 μg/ml respectively, which were significantly higher than those of PxB, 286
hBD1, hBD2, HNP-1 and magainin II for PxB-treated 52145-∆wcaK2 (2.6±0.7, 12.5±0.9, 7.0±0.5, 287
14±3.1 and 36.8 μg/ml, respectively, P < 0.05 for each comparison versus Kp52145 values [two-288
tailed t test]). Taken together, these observations suggest that PxB-induced resistance is in part 289
CPS-dependent but there are CPS-independent mechanisms operating as well. 290
Bacteria can modify the lipid A part of LPS by adding aminoarabinose, 291
phosphoethanolamine or palmitate to reduce the interaction of the peptides to the lipid A 292
(28,30,31,40,51). We speculated that the CPS-independent PxB-induced mechanism(s) of resistance 293
could involve changes in the lipid A structure. To explore this notion, we determined the structure 294
of lipid A extracted from Kp52145 after exposure to PxB by MALDI-TOF mass spectrometry (Fig 295
3). Lipid A from Kp52145 grown without PxB contained predominantly hexa-acylated species (m/z 296
1824) corresponding to two glucosamines, two phosphates, four 3-OH-C14, and two C14. Other peak 297
(m/z 1840) may represent a hexa-acylated lipid A containing two glucosamines, two phosphates, 298
four 3-OH-C14, one C14 and one C14:OH (hydroxymyristate). Minor species (m/z 1797) may 299
correspond to a hexa-acylated lipid A containing four 3-OH-C14, one C12 and one C14. Other minor 300
species detected were consistent with the addition of aminoarabinose (m/z 1955) to the hexa-301
acylated form (m/z 1824) or palmitate to the hexa-acylated species (m/z 1797 and 1824) hence 302
producing hepta-acylated lipid As (m/z 2036 and 2063) (Fig 3A). PxB induced an increase in the 303
relative abundance of the minor lipid A species containing aminoarabinose (m/z 1955) and 304
palmitate (m/z 2036 and 2063) (Fig 3B). Collectively, these results might indicate that exposure to 305
PxB increases K. pneumoniae lipid A modifications containing aminoarabinose and palmitate. 306
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 13 -
Persual of the literature shows that the products of ugd and pmrHFIJKLM (arnBCADTEF) 307
(hereafter pmrF operon) loci are required for the synthesis and addition of aminoarabinose to lipid 308
A. Ugd converts UDP-D-glucose into UDP-D-glucuronic acid which is next modified by pmrF 309
operon encoded enzymes to generate aminoarabinose (56). The gene encoding for the 310
acyltransferase pagP is required for the addition of palmitate to lipid A (31). To verify that these 311
loci were indeed implicated in PxB-triggered lipid A modifications with aminoarabinose and 312
palmitate, we analyzed the lipid A structure from 52145-ΔpmrF, 52145-ΔpagPGB and 52145-313
ΔpmrF-ΔpagPGB mutants. The three strains expressed the same amount of cell-bound CPS ( 65.6 ± 314
8 μg/104 CFU, 69.5 ± 7 μg/104 CFU, and 61.6 ± 9 μg/104 CFU respectively) as Kp52145 (64.4 ± 3 315
μg/104 CFU; P > 0.05 for each comparison versus Kp52145 value [two-tailed t test]). Results 316
shown in Figure 4, demonstrate that the pmrF mutant grown with PxB lacked lipid A species 317
containing aminoarabinose whereas the pagP mutant grown with PxB did not contain species 318
containing palmitate. 52145-ΔpmrF-ΔpagPGB lacked species containing aminoarabinose and 319
palmitate. It should be noted that lipid A species containing hydroxymyristate (C14:OH ; m/z 1840) 320
and palmitate (m/z 2036 and 2063) were not affected in 52145-ΔpmrF whereas lipid A species 321
containing hydroxymiristate and aminoarabinose (m/z 1955) were not affected in 52145-ΔpagPGB. 322
These findings led us to study whether PxB up-regulates the expression of ugd, pmrF operon 323
and pagP. To monitor transcription of these loci quantitatively, three transcriptional fusions were 324
constructed in which a promoterless lucFF was under the control of the loci promoter regions. The 325
fusions were introduced into Kp52145 and the luciferase activity was determined. PxB up-regulated 326
the expression of ugd::lucFF, pmrH::lucFF and pagP::lucFF transcriptional fusions (Fig 5) 327
thereby giving experimental support to our hypothesis. 328
329
Aminoarabinose and palmitate lipid A substitutions contribute to K. pneumoniae 330
antimicrobial peptide resistance. 331
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 14 -
We sought to determine the contribution of aminoarabinose and palmitate lipid A 332
substitutions to resistance against PxB and magainin II. 52145-ΔpmrF was more susceptible to PxB 333
than the wild-type strain but as susceptible as 52145-∆wcaK2, whereas 52145-∆wcaK2-ΔpmrF was 334
the most susceptible strain (Fig 6A). 52145-ΔpmrF was as resistant as Kp52145 to magainin II 335
hence indicating that the lipid A decoration with aminoarabinose is not implicated in the resistance 336
to this AP (Fig 6A). On the other hand, lipid A substitution with palmitate does not play any role in 337
PxB resistance but to other peptides such as magainin II (31). As expected, 52145-ΔpagPGB was as 338
resistant as Kp52145 to PxB (data not shown). However, 52145-ΔpagPGB was more susceptible 339
than Kp52145 to magainin II (Fig 6B). 52145-∆wcaK2-ΔpagPGB, lacking CPS and palmitate, was 340
the most susceptible strain to magainin II (Fig 6B). 341
We assessed the contribution of aminoarabinose or palmitate lipid A modifications to PxB-342
induced resistance to PxB and magainin II. The IC50 of PxB for PxB-treated Kp52145 was 4.5±0.8 343
μg/ml, which is significantly higher than those for 52145-ΔpmrF, 52145-∆wcaK2 and 52145-344
∆wcaK2-ΔpmrF (1.3±0.2, 2.6±0.5, and 0.3±0.2 μg/ml, respectively; P < 0.05 for each comparison 345
versus Kp52145 value [two-tailed t test]). PxB treatment also increased the resistance to magainin II 346
of 52145-ΔpagPGB (IC50 39.4±2.3 μg/ml) but the level was lower than that observed in the wild-347
type strain (56±4.3 μg/ml; P < 0.05 [two-tailed t test]) and similar to that of 52145-∆wcaK2 348
(41.3±4.2 μg/ml; P > 0.05 [two-tailed t test]). The lowest IC50 of magainin II was observed for 349
52145-∆wcaK2-ΔpagPGB (30.3±1.7 μg/ml). 350
Collectively, these data support the notion that CPS and the lipid A substitutions with 351
aminoarabinose and palmitate contribute to AP resistance in K. pneumoniae and also to PxB-352
induced cross resistance to APs. 353
354
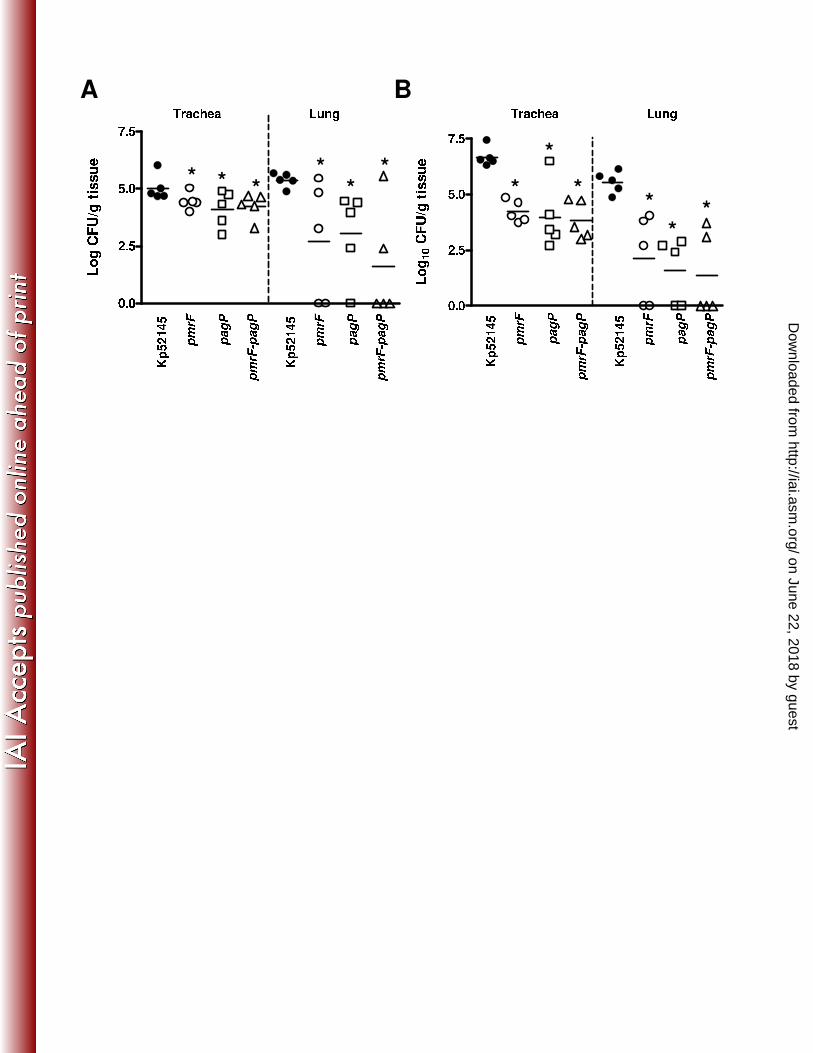
Virulence of K. pneumoniae lipid A mutans. 355
To determine the ability of 52145-ΔpmrF, 52145-ΔpagPGB and 52145-ΔpmrF-ΔpagPGB to 356
cause pneumonia, C57BL/6JOlaHsd mice were infected intranasally and 24 and 96 h post-infection 357
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 15 -
bacterial loads in trachea and lung homogenates were determined (Fig 7). At 24 h post-infection, all 358
strains colonized trachea and lungs although bacterial loads of mutant strains were lower than those 359
of the wild type in both organs (Fig 7A). A similar picture was observed at 96 h post infection (Fig 360
7B). Bacterial loads of mutants were not significantly different in either trachea or lungs at 24 and 361
96 h post infection (Fig 7). 362
363
Signaling networks controlling polymyxin B induced capsule and LPS lipid A modifications. 364
Having established that PxB up-regulated the expressions of cps, pmrF, ugd and pagP, we 365
sought to identify the regulatory architecture that mediates PxB-triggered up-regulation of them. 366
The Rcs (Regulator of capsule synthesis) phosphorelay system consists of three proteins; RcsC, 367
RcsD (also called YojN) and RcsB, the latter being a cytoplasmic response regulator (44). The Rcs 368
system fine tunes the expression of CPS in several Enterobacteriaceae and it mediates AP 369
resistance (15,65). PhoPQ and PmrAB two component systems mediate AP resistance by activating 370
loci leading to lipid A remodelling, including pagP, pmrF operon and ugd (25,27). In S. enterica, 371
the expression of pmrH and ugd is controlled by PmrAB whose activity can be modulated by the 372
PhoPQ-dependent PmrD connector protein at the post-transcriptional level (25,27,39). 373
To define the contribution of these systems to PxB-induced up-regulation of cps, pmrF, ugd 374
and pagP, we investigated the transcription of these loci in isogenic mutants with and without PxB 375
treatment. Basal levels of the cps transcriptional fusion were lower in 52145-ΔrcsB and 52145-376
ΔpmrAB than in Kp52145 (Fig 8A). PxB induced the fusion only in the 52145-ΔrcsB, 52145-377
ΔpmrD and 52145-ΔpmrAB backgrounds and to the same levels obtained in Kp52145 (Fig 8A). 378
PxB treatment up-regulated the expression of the pmrH transcriptional fusion in 52145-ΔrcsB, 379
52145-ΔphoQ, 52145-ΔpmrAB and 52145-ΔpmrD backgrounds although the levels obtained in the 380
52145-ΔrcsB background were significantly higher than those observed in the other strains (Fig 381
8B). A similar picture was observed for ugd::lucFF (Fig 8C). PxB did not up-regulate the 382
expression of pmrH and ugd fusions in the mutant lacking both phoQ and pmrAB (Fig 8B-C). PxB 383
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 16 -
up-regulated the pagP::lucFF fusion only in Kp52145, 52145-ΔrcsB, 52145-ΔpmrAB, and 52145-384
ΔpmrD (Fig 8D) and the levels found in the 52145-ΔrcsB background were the highest obtained 385
(Fig 8D). 386
To further sustain the role of PhoPQ in PxB-induced transcriptional effects, we analyzed 387
whether PxB up-regulates the expression of mgtA, whose expression is PhoPQ-dependent (60). As 388
expected, PxB up-regulated the expression of mgtA in Kp52145, 52145-ΔpmrAB and 52145-ΔpmrD 389
backgrounds to similar levels but not in 52145-ΔphoQ mutant (Fig 8E). Interestingly, mgtA 390
expression in the 52145-ΔrcsB background was significantly higher than those in the other strains 391
(Fig 8E). Finally, we tested whether PxB induced the expression of the pmrD connector in a 392
PhoPQ-dependent manner. Indeed, this was the case (Fig 8F). Furthermore, PxB-induced levels of 393
the pmrD fusion were similar in Kp52145 and 52145-ΔpmrAB backgrounds and the highest levels 394
were observed again in the 52145-ΔrcsB mutant background (Fig 8F). 395
In summary, these data indicate that PhoPQ is necessary for PxB-triggered induction of cps 396
and pagP. PxB induction of pmrH and ugd was only abolished in the double mutant phoQ-pmrAB 397
hence suggesting that both two component systems can promote PxB-induced lipid A modification 398
with aminoarabinose. Confirming this hypothesis, lipid A species containing aminoarabinose were 399
only absent in the PxB-treated phoQ-pmrAB double mutant background (supplemental Fig S1). 400
401
Cross-talk between Rcs and PhoPQ systems. 402
Considering that pmrH, ugd, pagP, mgtA and pmrD loci were overexpressed in the rcsB 403
mutant background and that PhoPQ regulates their expression, we hypothesized that the expression 404
of phoPQ could be up-regulated in the rcsB mutant background. To explore this, the expression of 405
phoP::lucFF was measured in different genetic backgrounds upon PxB treatment. Data shown in 406
Figure 9 revealed that phoP was overexpressed in 52145-ΔrcsB but down-regulated in 52145-407
ΔphoQ (Fig 9A). PxB up-regulated phoP transcription in Kp52145 and 52145-ΔrcsB backgrounds 408
being the highest levels those found in the latter. This was dependent on PhoPQ because phoP 409
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 17 -
transcription was not up-regulated in a double mutant lacking rcsB and phoQ (Fig 9A). phoP 410
expression was not affected in 52145-ΔpmrAB background (data not shown). Altogether, these data 411
gave experimental support to the hypothesis that phoPQ is up-regulated in the rcsB mutant 412
background. To further sustain this notion, we asked whether the expressions of pagP, mgtA and 413
pmrD are up-regulated in the 52145-ΔrcsB-phoQ mutant. As expected, this was not the case (Fig 414
9B-D). Furthermore, PxB did not up-regulate these loci in the 52145-ΔrcsB-phoQ background (Fig 415
9B-D) which is in good agreement with the findings showing that PhoPQ is necessary for their 416
PxB-mediated induction. Finally, we asked whether the up-regulation of pmrH and ugd obtained in 417
the rcsB mutant background was also dependent on PhoPQ. Indeed, the expression of pmrH and 418
ugd was not up-regulated in 52145-ΔrcsB-phoQ (Fig 9E-F). In sharp contrast with pagP, mgtA and 419
pmrD, the expressions of pmrH and ugd were still induced by PxB in 52145-ΔrcsB-phoQ (Fig 9E-420
F) which is consistent with our data showing that PmrAB also promotes PxB-induced up-regulation 421
of these loci (Fig 7). 422
We explored whether the expression of the Rcs system is affected in phoQ, pmrAB, phoQ-423
pmrAB and pmrD mutant backgrounds. To monitor transcription of the Rcs system, we analyzed the 424
expression of rcsD::lucFF and rcsC::lucFF transcriptional fusions. Basal levels of the rcsD fusion 425
were lower in 52145-ΔphoQ and 52145-ΔphoQ-pmrAB than those obtained in Kp52145, 52145-426
ΔpmrAB and 52145-ΔpmrD which were not significantly different between them (Fig 9G). A 427
similar picture was observed for the rcsC fusion (Fig 9H). rcsD transcription was down-regulated in 428
52145-ΔrcsB (Fig 9G) whereas the expression of the rcsC fusion was abolished in 52145-ΔrcsB 429
(Fig 9H). The former result is in good agreement with the auto regulation of the Rcs system 430
whereas the latter one is consistent with the fact that RcsB is essential for rcsC expression (44). PxB 431
treatment up-regulated the expression of rcsD however, PxB-induced rcsD expression was lower in 432
52145-ΔphoQ and 52145-ΔphoQ-pmrAB than those obtained in Kp52145, 52145-ΔpmrAB and 433
52145-ΔpmrD (Fig 9G). PxB did not induce the fusion in 52145-ΔrcsB (Fig 9G). Similar results 434
were observed when the expression of rcsC was analyzed upon PxB treatment (Fig 9H). 435
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 18 -
On the whole, these data support the notion that there is cross-talk between the Rcs and 436
PhoPQ systems. Whereas the Rcs system down-regulates phoP, the PhoPQ system promotes the 437
expressions of rcsD and rcsC. 438
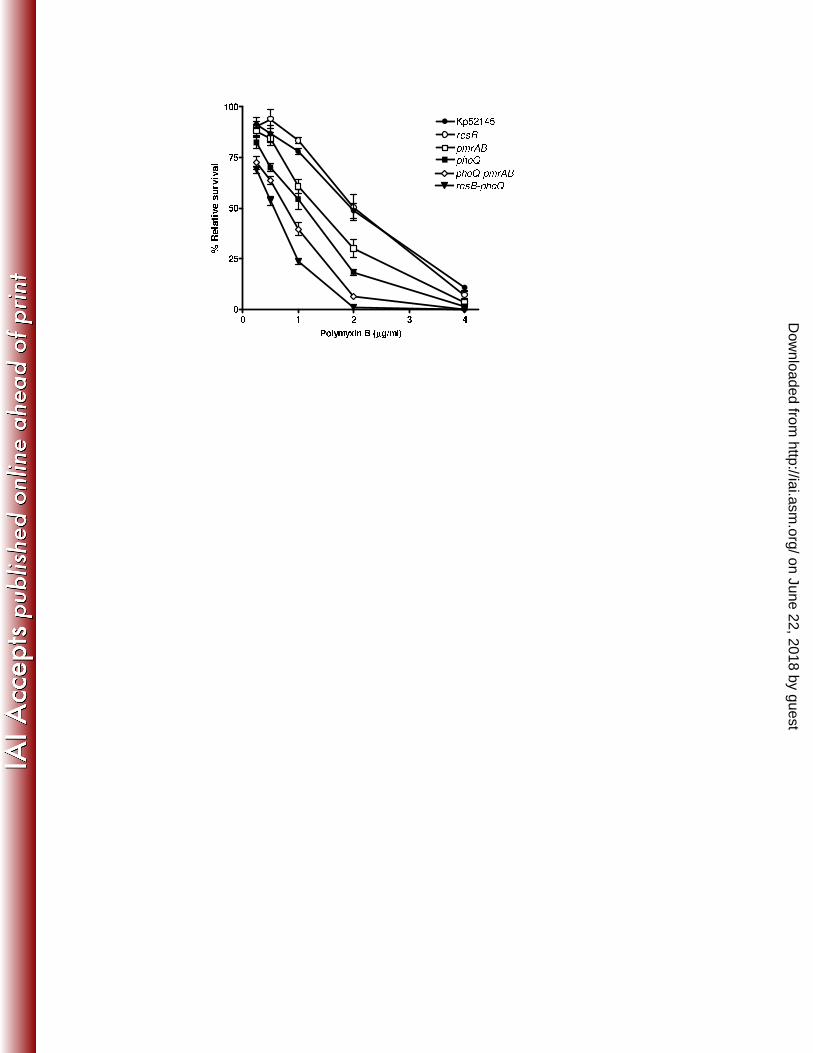
Finally, we tested the susceptibility to PxB of the three transduction systems. Results shown 439
in Figure 10 demonstrate that the most susceptible strain was 52145-ΔrcsB-phoQ (IC50; 0.42±0.2 440
μg/ml) followed by 52145-ΔphoQ-pmrAB (IC50; 0.77±0.1 μg/ml), the two single mutants 52145-441
ΔphoQ (IC50; 1.17±0.2 μg/ml) and 52145-ΔpmrAB (IC50; 1.25 ±0.3 μg/ml), and Kp52145 (IC50; 2.1 442
±0.3 μg/ml). 52145-ΔrcsB (IC50; 2.3 ±0.4 μg/ml) was as susceptible as the wild type. 443
444
DISCUSSION 445
In the present study, we provide new insights into how a bacterial pathogen activates 446
countermeasures to fight against APs. Our findings revealed that brief treatment of a virulent isolate 447
of K. pneumoniae with PxB induces cross-resistance to APs found in humans as well as magainin II 448
and PxB. Mechanistically, PxB triggers changes in K. pneumoniae surface which contribute to AP 449
resistance and PxB-induced cross resistance. Finally, we demonstrate that lipid A modifications are 450
important for K. pneumoniae survival in the airways. 451
Our data showed that PxB induced cross-resistance to APs not structurally related (32,62) 452
thereby indicating that PxB-triggered resistance is not specific for the compound used. Since APs 453
share the initial electrostatic interaction with the anionic bacterial surface we hypothesized that PxB 454
treatment should affect the bacterial surface. Indeed, PxB treatment up-regulated the expression of 455
the cps operon and the loci required to modify the lipid A with aminoarabinose and palmitate with a 456
concomitant increase in CPS and lipid A species containing such modifications. Moreover, these 457
surface changes were linked to AP resistance. Previous data (10,43), further confirmed here, had 458
shown that K. pneumoniae CPS mediates resistance to several APs. In this work, we also 459
demonstrate that lipid A modifications with aminoarabinose and palmitate are required for AP 460
resistance. It should be noted that these Klebsiella countermeasures are not redundant since double 461
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 19 -
mutants lacking CPS and lipid A modifications were more susceptible to APs than the single 462
mutants. Although here we have just considered surface changes we do not rule out that other 463
bacterial systems could be affected as well. In fact, we put forward the notion that upon challenge 464
with PxB, or other APs such as defensins, K. pneumoniae may alter global gene expression. It is 465
tempting to speculate that these bacterial global changes could be a “molecular pattern” devoted to 466
counteract the innate immune system including the activation of inflammatory responses and the 467
microbicidal action of professional phagocytes (macrophages and neutrophils). Studies are on-going 468
to confirm this hypothesis. 469
Lipid A analysis revealed the presence of a species (m/z 1840) consistent with the presence 470
of hydroxymyristate (C14:OH). This species has been previously reported for K. pneumoniae (11) and 471
S. enterica serovar typhimurium (23). For the latter, it has been identified the dioxygenase 472
responsible for 2-hydroxylation, named LpxO (23). This enzyme generates 2-hydroxymyristate by 473
hydroxylation of the myristate fatty acid transferred to lipid A by the acyltransferase MsbB/LpxM 474
(23). In silico analysis of the available K. pneumoniae genomes revealed that this pathogen may 475
encode an orthologue of LpxO. Studies are ongoing to characterize K. pneumoniae LpxO and 476
whether this lipid A modification plays any role in the resistance to APs. A remaining question is to 477
explain at the molecular level the species (m/z 1744) found only in those strains treated with PxB. 478
This species is consistent with elimination of one phosphate from the molecular ion m/z 1824. 479
Among other possibilities, it can be speculated that PxB treatment activates a lipid A phosphatase. 480
In fact, lipid A dephosphorylation mediates AP resistance in Phorphyromonas, Rhizobium, 481
Francisella and Helicobacter (12,38,61,64). Future studies will aim to identify this putative K. 482
pneumoniae lipid A phosphatase. 483
Our analysis of the regulatory architecture governing PxB-induced changes revealed that the 484
two-component system PhoPQ is necessary for PxB induction of cps, pagP, mgtA and pmrD. These 485
findings further support the notion that brief treatment with APs activates the PhoPQ regulon as 486
first reported by Bader and co-workers in S. enterica serovar typhimurium (4). However, the fact 487
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 20 -
that PxB treatment increased the expressions of rcsC, rcsD, pmrH and ugd in the phoQ mutant 488
suggests that PhoPQ is not the only sensor/regulator governing PxB-induced AP resistance in K. 489
pneumoniae. Indeed, our results demonstrate that PmrAB also transduces the PxB-dependent 490
regulatory signal. Thus, only in the double mutant phoQ-pmrAB background, PxB did not up-491
regulate the expressions of pmrH and ugd which is consistent with the presence of PhoP and PmrA 492
boxes in the promoter regions of both loci (47). This is in contrast to what happens in Salmonella 493
since, under the conditions used in the present study, PxB-dependent up-regulation of pmrH and 494
ugd is dependent on PmrAB via activation of PhoPQ and PmrD (27,28,30,39). Interestingly, PxB 495
treatment activated the Rcs system as detected by the up-regulation of the rcsC and rcsD 496
transcriptional fusions. Despite that the levels of both fusions were 20% lower in phoQ and phoQ-497
pmrAB mutants than in Kp52145, PxB still increased the expression of rcsC and rcsD hence 498
suggesting that, at least, an additional regulatory system may govern PxB-induced changes. Our 499
data suggest that indeed the Rcs system could be this other system since PxB did not longer induce 500
the rcsD fusion in the rcsB mutant. Furthermore, our observation that the most susceptible strain to 501
PxB was the double mutant rcsB-phoQ suggests that there are Rcs-dependent responses implicated 502
in the resistance to APs. This is not in contradiction with the fact that the rcsB mutant was as 503
susceptible as the wild-type strain to PxB because we have demonstrated that PhoPQ-dependent 504
responses were up-regulated in this genetic background. Therefore, it is tempting to conclude that 505
these putative Rcs-dependent AP countermeasures play a role only in the absence of the PhoPQ-506
dependent ones. 507
An important issue is to understand how these signaling systems sense APs to activate a 508
transcriptional program. It has been proposed that Salmonella PhoQ binds APs in an acidic patch of 509
the periplasmic domain which leads to PhoQ auto-phosphorylation and the subsequent 510
phosphotransfer to PhoP (5,54,55). K. pneumoniae PhoQ also contains this acidic patch and 511
therefore a similar mechanism of PhoQ activation by APs might be expected. At present, we can 512
only speculate on how the PmrAB and Rcs systems are activated by APs. To date no structural 513
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 21 -
study has analyzed in-depth PmrB proteins though the activation of Salmonella PmrB has been 514
linked to the binding of cations to a periplasmic domain (27). Alternatively, activation of the 515
membrane sensors could result from outer membrane perturbations and not involve a direct 516
interaction with APs. Considering that all APs disorganize the outer membrane (62), it would be 517
logical that Gram-negative pathogens have evolved means to detect membrane integrity through the 518
use of membrane located proteins. In support of this model, it has been shown recently that different 519
APs activate the Rcs system through increased accessibility of the RcsF protein to the inner 520
membrane or periplasm (18). It is possible that APs may activate each signaling system in a 521
different way which will give Gram-negative bacteria the opportunity to integrate different signals 522
to promote phenotypic changes leading to AP resistance. This could also explain why the 523
differences between the wild type and the single mutants are not dramatic since the response is 524
mediated by a collective effort of, at least, three signaling systems. 525
Here, we demonstrate, for the first time using a pneumonia mouse model, that lipid A 526
modifications are important for bacterial survival in the lung. Previously we did demonstrate that 527
CPS mediates resistance against APs (10,43) and it is known that CPS is an important Klebsiella 528
virulence factor (13). Collectively, it can be postulated that there is a correlation between resistance 529
to APs and the ability to cause pneumonia. There are several APs in the airway liquid including 530
lysozyme, lactoferrin, α-defensins, β-defensins and cathelicidins (1). Therefore, the mutants’ 531
susceptibility to APs could explain the decrease bacterial loads of these strains in airways. 532
However, the in vivo scenario is complex and the final outcome of pneumonia is the combination of 533
the action of antimicrobial factors (among others complement and APs) and several types of cells 534
including alveolar macrophages, epithelial cells and neutrophils. It should be noted that cytokines 535
and chemokines released by epithelial cells do up-regulate the expression of APs and also increase 536
the bactericidal activity of professional phagocytes. Studies are ongoing to determine whether lipid 537
A modifications play any role in the interplay of K. pneumoniae with airway cells. 538
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 22 -
Polymyxins are considered the “last hope” to treat infections caused by multi-drug resistant 539
Gram-negative bacteria including K. pneumoniae. Our results demonstrate that exposure to PxB 540
induces cross-resistance to PxB itself but also to APs present in the airways. Interestingly, results 541
from our laboratory indicate that K. pneumoniae prevents the expression of APs by airway cells 542
(48). Collectively, these findings are consistent with a scenario in which the setting of pneumonia 543
by K. pneumoniae will be facilitated by, on one hand, preventing the expression of APs and, on the 544
other hand, activating countermeasures against them. In turn, we put forward the idea that 545
therapeutic strategies directed to prevent the activation of this program could be a new approach 546
worthy exploring to facilitate the clearance of the pathogen from the airways. 547
548
ACKNOWLEDGEMENTS 549
We are grateful to members of Bengoechea lab for helpful discussions and Christian Frank for 550
critically reading the manuscript. This work has been funded by grants from Fondo de Investigación 551
Sanitaria (PI06/1629), and Biomedicine Program (SAF2009-07885) from Ministerio de Ciencia e 552
Innovación (Spain) to J.A.B. CIBERES is an initiative from Instituto de Salud Carlos III. 553
554
Reference List 555 556
1. Agerberth, B., J. Grunewald, E. Castanos-Velez, B. Olsson, H. Jornvall, H. Wigzell, A. 557 Eklund, and G. H. Gudmundsson. 1999. Antibacterial components in bronchoalveolar 558 lavage fluid from healthy individuals and sarcoidosis patients. Am. J Respir. Crit Care Med. 559 160:283-290. 560
2. Appleyard, R. K. 1954. Segregation of new lysogenic types during growth of doubly 561 lysogenic strain derived from Escherichia coli K12. Genetics 39:440-452. 562
3. Aussel, L., H. Thérisod, D. Karibian, M. B. Perry, M. Bruneteau, and M. Caroff. 2000. 563 Novel variation of lipid A structures in strains of different Yersinia species. FEBS Letters 564 465:87-92. 565
4. Bader, M. W., W. W. Navarre, W. Shiau, H. Nikaido, J. G. Frye, M. Mcclelland, F. C. 566 Fang, and S. I. Miller. 2003. Regulation of Salmonella typhimurium virulence gene 567 expression by cationic antimicrobial peptides. Mol. Microbiol. 50:219-230. 568
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 23 -
5. Bader, M. W., S. Sanowar, M. E. Daley, A. R. Schneider, U. Cho, W. Xu, R. E. Klevit, 569 M. H. Le, and S. I. Miller. 2005. Recognition of antimicrobial peptides by a bacterial 570 sensor kinase. Cell 122:461-472. 571
6. Behlau, I. and S. I. Miller. 1993. A PhoP-repressed gene promotes Salmonella 572 typhimurium invasion of epithelial cells. J. Bacteriol. 175:4475-4484. 573
7. Bitter, T. and H. M. Muir. 1962. A modified uronic acid carbazole reaction. Anal. 574 Biochem. 4:330-334. 575
8. Brogden, K. A. 2005. Antimicrobial peptides: pore formers or metabolic inhibitors in 576 bacteria? Nat. Rev. Microbiol. 3:238-250. 577
9. Byrappa, S., D. K. Gavin, and K. C. Gupta. 1995. A highly efficient procedure for site-578 specific mutagenesis of full- length plasmids using Vent DNA polymerase. Genome 579 Research 5:404-407. 580
10. Campos, M. A., M. A. Vargas, V. Regueiro, C. M. Llompart, S. Alberti, and J. A. 581 Bengoechea. 2004. Capsule polysaccharide mediates bacterial resistance to antimicrobial 582 peptides. Infect. Immun. 72:7107-7114. 583
11. Clements, A., D. Tull, A. W. Jenney, J. L. Farn, S. H. Kim, R. E. Bishop, J. B. McPhee, 584 R. E. Hancock, E. L. Hartland, M. J. Pearse, O. L. Wijburg, D. C. Jackson, M. J. 585 McConville, and R. A. Strugnell. 2007. Secondary acylation of Klebsiella pneumoniae 586 lipopolysaccharide contributes to sensitivity to antibacterial peptides. J. Biol. Chem. 587 282:15569-15577. 588
12. Coats, S. R., J. W. Jones, C. T. Do, P. H. Braham, B. W. Bainbridge, T. T. To, D. R. 589 Goodlett, R. K. Ernst, and R. P. Darveau. 2009. Human Toll-like receptor 4 responses to 590 P. gingivalis are regulated by lipid A 1- and 4'-phosphatase activities. Cell Microbiol. 591 11:1587-1599. 592
13. Cortes, G., N. Borrell, B. de Astorza, C. Gomez, J. Sauleda, and S. Alberti. 2002. 593 Molecular analysis of the contribution of the capsular polysaccharide and the 594 lipopolysaccharide O side chain to the virulence of Klebsiella pneumoniae in a murine 595 model of pneumonia. Infect. Immun. 70:2583-2590. 596
14. Datsenko, K. A. and B. L. Wanner. 2000. One-step inactivation of chromosomal genes in 597 Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. U. S. A 97:6640-6645. 598
15. Detweiler, C. S., D. M. Monack, I. E. Brodsky, H. Mathew, and S. Falkow. 2003. virK, 599 somA and rcsC are important for systemic Salmonella enterica serovar Typhimurium 600 infection and cationic peptide resistance. Mol. Microbiol. 48:385-400. 601
16. El, H. A., A. Tirsoaga, A. Novikov, A. Hussein, and M. Caroff. 2005. Microextraction of 602 bacterial lipid A: easy and rapid method for mass spectrometric characterization. J. Lipid 603 Res. 46:1773-1778. 604
17. Evans, M. E., D. J. Feola, and R. P. Rapp. 1999. Polymyxin B sulfate and colistin: old 605 antibiotics for emerging multiresistant gram-negative bacteria. Ann. Pharmacother. 33:960-606 967. 607
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 24 -
18. Farris, C., S. Sanowar, M. W. Bader, R. Pfuetzner, and S. I. Miller. 2010. Antimicrobial 608 peptides activate the Rcs regulon through the outer membrane lipoprotein RcsF. J. Bacteriol. 609 192:4894-4903. 610
19. Favre, D. and J. F. Viret. 2000. Gene replacement in gram-negative bacteria: the 611 pMAKSAC vectors. Biotechniques 28:198-200, 202, 204. 612
20. Fields, P. I., E. A. Groisman, and F. Heffron. 1989. A Salmonella locus that controls 613 resistance to microbicidal proteins from phagocytic cells. Science 243:1059-1062. 614
21. Fields, P. I., R. V. Swanson, C. G. Haidaris, and F. Heffron. 1986. Mutants of 615 Salmonella typhimurium that cannot survive within the macrophage are avirulent. Proc. 616 Natl. Acad. Sci. U. S. A 83:5189-5193. 617
22. Ganz, T. 1987. Extracellular release of antimicrobial defensins by human 618 polymorphonuclear leukocytes. Infect. Immun. 55:568-571. 619
23. Gibbons, H. S., S. Lin, R. J. Cotter, and C. R. Raetz. 2000. Oxygen requirement for the 620 biosynthesis of the S-2-hydroxymyristate moiety in Salmonella typhimurium lipid A. 621 Function of LpxO, A new Fe2+/alpha-ketoglutarate-dependent dioxygenase homologue. J. 622 Biol. Chem. 275:32940-32949. 623
24. Gootz, T. D. 2010. The global problem of antibiotic resistance. Crit Rev. Immunol. 30:79-624 93. 625
25. Groisman, E. A. 2001. The pleiotropic two-component regulatory system PhoP-PhoQ. J. 626 Bacteriol. 183:1835-1842. 627
26. Groisman, E. A., C. Parralopez, M. Salcedo, C. J. Lipps, and F. Heffron. 1992. 628 Resistance to Host Antimicrobial Peptides Is Necessary for Salmonella Virulence. Proc. 629 Natl. Acad. Sci. U. S. A 89:11939-11943. 630
27. Gunn, J. S. 2008. The Salmonella PmrAB regulon: lipopolysaccharide modifications, 631 antimicrobial peptide resistance and more. Trends Microbiol. 16:284-290. 632
28. Gunn, J. S., K. B. Lim, J. Krueger, K. Kim, L. Guo, M. Hackett, and S. I. Miller. 1998. 633 PmrA-PmrB-regulated genes necessary for 4-aminoarabinose lipid A modification and 634 polymyxin resistance. Mol. Microbiol. 27:1171-1182. 635
29. Gunn, J. S. and S. I. Miller. 1996. Pho-PhoQ activates transcription of pmrAB, encoding a 636 two-component regulatory system involved in Salmonella typhimurium antimicrobial 637 peptide resistance. J. Bacteriol 178:6857-6864. 638
30. Gunn, J. S., S. S. Ryan, J. C. Van Velkinburgh, R. K. Ernst, and S. I. Miller. 2000. 639 Genetic and functional analysis of a PmrA-PmrB-regulated locus necessary for 640 lipopolysaccharide modification, antimicrobial peptide resistance, and oral virulence of 641 Salmonella enterica serovar typhimurium. Infect Immun 68:6139-6146. 642
31. Guo, L., K. B. Lim, C. M. Poduje, M. Daniel, J. S. Gunn, M. Hackett, and S. I. Miller. 643 1998. Lipid A acylation and bacterial resistance against vertebrate antimicrobial peptides. 644 Cell 95:189-98. 645
32. Hancock, R. E. W. and D. S. Chapple. 1999. Peptide antibiotics. Antimicrob Agents 646 Chemother 43:1317-1323. 647
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 25 -
33. Harder, J., J. Bartels, E. Christophers, and J. M. Schroder. 2001. Isolation and 648 characterization of human beta -defensin-3, a novel human inducible peptide antibiotic. J 649 Biol. Chem. 276:5707-5713. 650
34. Harder, J., U. Meyer-Hoffert, L. M. Teran, L. Schwichtenberg, J. Bartels, S. Maune, 651 and J. M. Schroder. 2000. Mucoid Pseudomonas aeruginosa, TNF-alpha, and IL-1beta, 652 but not IL-6, induce human beta-defensin-2 in respiratory epithelia. Am. J Respir. Cell Mol 653 Biol. 22:714-721. 654
35. Hermsen, E. D., C. J. Sullivan, and J. C. Rotschafer. 2003. Polymyxins: pharmacology, 655 pharmacokinetics, pharmacodynamics, and clinical applications. Infect. Dis. Clin. North 656 Am. 17:545-562. 657
36. Hiratsuka, T., M. Nakazato, Y. Date, J. Ashitani, T. Minematsu, N. Chino, and S. 658 Matsukura. 1998. Identification of human beta-defensin-2 in respiratory tract and plasma 659 and its increase in bacterial pneumonia. Biochem Biophys. Res. Commun. 249:943-947. 660
37. Hoang, T. T., R. R. Karkhoff-Schweizer, A. J. Kutchma, and H. P. Schweizer. 1998. A 661 broad-host-range Flp-FRT recombination system for site-specific excision of 662 chromosomally-located DNA sequences: application for isolation of unmarked 663 Pseudomonas aeruginosa mutants. Gene 212:77-86. 664
38. Karbarz, M. J., S. R. Kalb, R. J. Cotter, and C. R. Raetz. 2003. Expression cloning and 665 biochemical characterization of a Rhizobium leguminosarum lipid A 1-phosphatase. J. Biol. 666 Chem. 278:39269-39279. 667
39. Kox, L. F., M. M. Wösten, and E. A. Groisman. 2000. A small protein that mediates the 668 activation of a two-component system by another two-component system. EMBO J. 669 19:1861-1872. 670
40. Lee, H., F. F. Hsu, J. Turk, and E. A. Groisman. 2004. The PmrA-regulated pmrC gene 671 mediates phosphoethanolamine modification of lipid A and polymyxin resistance in 672 Salmonella enterica. J. Bacteriol. 186:4124-4133. 673
41. Lindner, B. 2000. Matrix-assisted laser desorption/ionization time-of-flight mass 674 spectrometry of lipopolysaccharides. Methods Mol. Biol. 145:311-325. 675
42. Link, A. J., D. Phillips, and G. M. Church. 1997. Methods for generating precise 676 deletions and insertions in the genome of wild-type Escherichia coli: application to open 677 reading frame characterization. J. Bacteriol. 179:6228-6237. 678
43. Llobet, E., J. M. Tomas, and J. A. Bengoechea. 2008. Capsule polysaccharide is a 679 bacterial decoy for antimicrobial peptides. Microbiology 154:3877-3886. 680
44. Majdalani, N. and S. Gottesman. 2005. The Rcs phosphorelay: a complex signal 681 transduction system. Annu. Rev. Microbiol. 59:379-405. 682
45. Matsuzaki, K. 1998. Magainins as paradigm for the mode of action of pore forming 683 polypeptides. Biochim. Biophys. Acta 1376:391-400. 684
46. McCray, P. B., Jr. and L. Bentley. 1997. Human airway epithelia express a beta-defensin. 685 Am. J Respir. Cell Mol Biol. 16:343-349. 686
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 26 -
47. Mitrophanov, A. Y., M. W. Jewett, T. J. Hadley, and E. A. Groisman. 2008. Evolution 687 and dynamics of regulatory architectures controlling polymyxin B resistance in enteric 688 bacteria. PLoS. Genet. 4:e1000233. 689
48. Moranta, D., V. Regueiro, C. March, E. Llobet, J. Margareto, E. Larrate, J. 690 Garmendia, and J. A. Bengoechea. 2010. Klebsiella pneumoniae capsule polysaccharide 691 impedes the expression of β-defensins by airway epithelial cells. Infect. Immun. 78:1135-692 1146. 693
49. Nassif, X., J. M. Fournier, J. Arondel, and P. J. Sansonetti. 1989. Mucoid phenotype of 694 Klebsiella pneumoniae is a plasmid-encoded virulence factor. Infect Immun 57:546-552. 695
50. Nicolas, P. and A. Mor. 1995. Peptides as weapons against microorganisms in the chemical 696 defense system of vertebrates. Annu Rev Microbiol 49:277-304. 697
51. Nizet, V. 2006. Antimicrobial peptide resistance mechanisms of human bacterial pathogens. 698 Curr. Issues Mol. Biol. 8:11-26. 699
52. Oertelt, C., B. Lindner, M. Skurnik, and O. Holst. 2001. Isolation and structural 700 characterization of an R-form lipopolysaccharide from Yersinia enterocolitica serotype O:8. 701 Eur. J. Biochem. 268:554-564. 702
53. Perez-Gutierrez, C., E. Llobet, C. M. Llompart, M. Reines, and J. A. Bengoechea. 703 2010. Role of lipid A acylation in Yersinia enterocolitica virulence. Infect. Immun. 704 78:2768-2781. 705
54. Prost, L. R., M. E. Daley, M. W. Bader, R. E. Klevit, and S. I. Miller. 2008. The PhoQ 706 histidine kinases of Salmonella and Pseudomonas spp. are structurally and functionally 707 different: evidence that pH and antimicrobial peptide sensing contribute to mammalian 708 pathogenesis. Mol. Microbiol. 69:503-519. 709
55. Prost, L. R., M. E. Daley, S. Le, V, M. W. Bader, M. H. Le, R. E. Klevit, and S. I. 710 Miller. 2007. Activation of the bacterial sensor kinase PhoQ by acidic pH. Mol. Cell 711 26:165-174. 712
56. Raetz, C. R., C. M. Reynolds, M. S. Trent, and R. E. Bishop. 2007. Lipid A modification 713 systems in gram-negative bacteria. Annu. Rev. Biochem. 76:295-329. 714
57. Rahn, A. and C. Whitfield. 2003. Transcriptional organization and regulation of the 715 Escherichia coli K30 group 1 capsule biosynthesis (cps) gene cluster. Mol Microbiol 716 47:1045-1060. 717
58. Rebeil, R., R. K. Ernst, B. B. Gowen, S. I. Miller, and B. J. Hinnebusch. 2004. Variation 718 in lipid A structure in the pathogenic yersiniae. Mol. Microbiol. 52:1363-1373. 719
59. Singh, P. K., H. P. Jia, K. Wiles, J. Hesselberth, L. Liu, B. A. Conway, E. P. 720 Greenberg, E. V. Valore, M. J. Welsh, T. Ganz, B. F. Tack, and P. B. McCray, Jr. 721 1998. Production of beta-defensins by human airway epithelia. Proc Natl Acad Sci U S A 722 95:14961-14966. 723
60. Soncini, F. C., V. E. Garcia, F. Solomon, and E. A. Groisman. 1996. Molecular basis of 724 the magnesium deprivation response in Salmonella typhimurium: identification of PhoP-725 regulated genes. J. Bacteriol. 178:5092-5099. 726
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 27 -
61. Tran, A. X., J. D. Whittimore, P. B. Wyrick, S. C. McGrath, R. J. Cotter, and M. S. 727 Trent. 2006. The lipid A 1-phosphatase of Helicobacter pylori is required for resistance to 728 the antimicrobial peptide polymyxin. J. Bacteriol. 188:4531-4541. 729
62. Vaara, M. 1992. Agents that increase the permeability of the outer membrane. Microbiol 730 Rev 56:395-411. 731
63. Vaara, M. 2010. Polymyxins and their novel derivatives. Curr. Opin. Microbiol. 13:574-732 581. 733
64. Wang, X., S. C. McGrath, R. J. Cotter, and C. R. Raetz. 2006. Expression cloning and 734 periplasmic orientation of the Francisella novicida lipid A 4'-phosphatase LpxF. J. Biol. 735 Chem. 281:9321-9330. 736
65. Wehland, M. and F. Bernhard. 2000. The RcsAB box. Characterization of a new operator 737 essential for the regulation of exopolysaccharide biosynthesis in enteric bacteria. J. Biol. 738 Chem. 275:7013-7020. 739
740
741 742
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 28 -
FIGURE LEGENDS 743
Figure 1. Exposure of K. pneumoniae 52145 to polymyxin B increases the resistance to 744
antimicrobial peptides. After polymyxin B-treatment bacteria were washed and the susceptibility 745
to (A) polymyxin B; (B) β-defensin 1, (C) β-defensin 2; (D) HNP-1; and (E) magainin II tested by 746
the survival assay. 747
Each point represents the mean and standard deviation of eight samples from four independently 748
grown batches of bacteria and significant survival differences (P < 0.05; two-tailed t test) between 749
bacteria pre-treated with polymyxin B (black symbols) and non-treated bacteria (white symbols) are 750
indicated by asterisks. 751
752
Figure 2. Treatment of K. pneumoniae 52145 cps mutant with polymyxin B increases the 753
resistance to antimicrobial peptides. (A) Analysis of the expression of the cps operon by 754
Kp52145 carrying the fusion cps::lucFF. The strain was treated with polymyxin B for 1 h (black 755
bar) and non-treated (white bar). Data are presented as mean ± SD (n = 6). *, results are 756
significantly different (P < 0.05; two-tailed t test) from the results for non-treated bacteria. (B-F) 757
After 1 h treatment of 52145-∆wcaK2 with polymyxin B, bacteria were washed and exposed to 758
different concentrations of (B) polymyxin B; (C) β-defensin 1, (D) β-defensin 2; (E) HNP-1; and 759
(F) magainin II. Each point represents the mean and standard deviation of eight samples from four 760
independently grown batches of bacteria and significant survival differences (P < 0.05; two-tailed t 761
test) between bacteria pre-treated with polymyxin B (black symbols) and non-treated bacteria 762
(white symbols) are indicated by asterisks. 763
764
Figure 3. Exposure of K. pneumoniae 52145 to polymyxin B affects the lipid A structure. 765
Negative ion MALDI-TOF mass spectrometry spectra of lipid A isolated from Kp52145 which was 766
treated with 65 ng/ml polymyxin B for 12 h (B) or not (A). The results in panels are representative 767
of three independent lipid A extractions. (C) Proposed structures corresponding to major peaks and 768
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 29 -
acyl group positions follow previously reported structures for Klebsiella (11) and other Gram-769
negative bacteria 770
771
Figure 4. Lipid A analysis from K. pneumoniae lipid A mutants. Negative ion MALDI-TOF 772
mass spectrometry spectra of lipid A isolated from the indicated K. pneumoniae strains which were 773
treated with 65 ng/ml polymyxin B for 12 h (B, D, F) or not (A, C, E). The results in all panels are 774
representative of three independent lipid A extractions. 775
776
Figure 5. Polymyxin B induces the expression of K. pneumoniae 52145 ugd, pmrF and pagP 777
loci. Analysis of the expression of the loci implicated in lipid A remodeling by measuring luciferase 778
activity of Kp52145 carrying ugd::lucFF, pmrF::lucFF or pagP::lucFF transcriptional fusions 779
which were treated with polymyxin B for 1 h (black bars) or non-treated (white bars). Data are 780
presented as mean ± SD (n = 5). *, results are significantly different (P < 0.05; two-tailed t test) 781
from the results for non-treated bacteria. 782
783
Figure 6. Roles of K. pneumoniae 52145 capsule and lipid A modifications on the resistance to 784
antimicrobial peptides. (A) pmrF mutants were exposed to different concentrations of polymyxin 785
B; and magainin II. (B) pagP mutants were exposed to different concentrations of magainin II. Each 786
point represents the mean and standard deviation of eight samples from four independently grown 787
batches of bacteria. 788
Symbols: , Kp52145; , 52145-∆wcaK2; , 52145-∆pmrF; , 52145-∆wcaK2-ΔpmrF; , 789
52145-ΔpagPGB; , 52145-∆wcaK2-ΔpagPGB. 790
791
Figure 7. Virulence of K. pneumoniae 52145 lipid A mutants. Bacterial counts in mouse organs 792
at 24 h (A) or 96 h (B) post infection. Mice were infected intranasally with a bacterial mixture 793
containing 4.1 x 104 bacteria of wild type (Kp52145, ); 4.5 x 104 bacteria of 52145-ΔpmrF (pmrF, 794
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 30 -
); 5.1 x 104 bacteria of 52145-ΔpagPGB (pagP, ) and 4.8 x 104 bacteria of 52145-ΔpmrF-795
ΔpagPGB (pmrF-pagP, ), respectively. Results were reported as log CFU per gram of tissue 796
(Log CFU/g). *, results are significantly different (P < 0.05; two-tailed t test) from the results for 797
Kp52145. 798
799
Figure 8. K. pneumoniae PhoPQ and PmrAB two component systems control polymyxin B-800
induced transcriptional changes. Analysis of the expression of cps, pmrH, ugd, pagP, mgtA and 801
pmrD loci by Kp52145 (WT), 52145-ΔrcsB (rcsB), 52145-ΔphoQGB (phoQ), 52145-ΔpmrAB 802
(pmrAB), 52145-ΔpmrD (pmrD) and 52145-ΔphoQGB-ΔpmrAB (phoQ-pmrAB) carrying the 803
transcriptional fusions cps::lucFF (A); pmrF::lucFF (B); ugd::lucFF (C); pagP::lucFF (D); 804
mgtA::lucFF (E); and pmrD::lucFF (F) treated with polymyxin B for 1 h (black bars) or non-805
treated (white bars). Data are presented as mean ± SD (n = 3). *, results are significantly different 806
(P < 0.05; two-tailed t test) from the results for non-treated bacteria. , results are significantly 807
different (P < 0.05; two-tailed t test) from the results for Kp52145 treated in the same manner. 808
809
Figure 9. There is cross-talk between Rcs and PhoPQ systems in K. pneumoniae 52145. 810
Analysis of the expression of phoP, pagP, mgtA and pmrD, pmrH, ugd, rcsD and rcsC loci by 811
Kp52145 (WT), 52145-ΔrcsB (rcsB), 52145-ΔphoQGB (phoQ), 52145-ΔrcsB−ΔphoQGB (rcsB-812
phoQ) carrying the transcriptional fusions phoP::lucFF (A); pagP::lucFF (B); mgtA::lucFF (C); 813
pmrD::lucFF (D); pmrH::lucFF (E); ugd::lucFF (F); rcsD::lucFF (G); and rcsC::lucFF (H) 814
treated with polymyxin B for 1 h (black bars) or non-treated (white bars). Data are presented as 815
mean ± SD (n = 3). *, results are significantly different (P < 0.05; two-tailed t test) from the results 816
for non-treated bacteria. , results are significantly different (P < 0.05; two-tailed t test) from the 817
results for Kp52145 treated in the same manner. 818
819
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 31 -
Figure 10. Role of K. pneumoniae Rcs, PhoPQ and PmrAB systems in bacterial susceptibility 820
to polymyxin B. Wild type (Kp52145), 52145-ΔrcsB (rcsB), 52145-ΔphoQGB (phoQ), 52145-821
ΔpmrAB (pmrAB), 52145-ΔphoQGB-ΔpmrAB (phoQ-pmrAB), and 52145-ΔrcsB−ΔphoQGB (rcsB-822
phoQ) were exposed to different concentrations of polymyxin B. Each point represents the mean 823
and standard deviation of eight samples from four independently grown batches of bacteria. 824
825
826
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 32 -
Table 1. Bacterial strains and plasmids used in this study. 827 828 829 Bacterial strains and plasmids Genotype or comments Source or
references Strains Escherichia coli C600 Thi, thr, leuB, tonA, lacy, supE (2) XL-1 Blue recA1, endA1 gyrA96 thi-1 hsdR17
supE44
S17-1λpir recA thi pro hsdR-M+ RP4::2-Tc::Mu::Km, Tn7λ pir
Klebsiella pneumoniae Kp52145 clinical isolate (serotype O1:K2), RifR (13,49) 52145-ΔwcaK2 Kp52145, ΔwcaK2 , RifR; the wcaK2 gene
inactivated, no CPS expression (43)
52145-ΔphoQGB Kp52145, ΔphoQ::Km-GenBlock , RifR,KmR;the phoQ gene inactivated
This study
52145-ΔpmrAB Kp52145, ΔpmrAB , RifR, the pmrAB genes inactivated
This study
52145-ΔrcsB Kp52145, ΔrcsB, RifR, the rcsB gene inactivated
This study
52145-ΔpmrD Kp52145, ΔpmrD , RifR, the pmrD gene inactivated
This study
52145-ΔpmrF Kp52145, ΔpmrF , RifR, the pmrF gene inactivated
This study
52145-ΔpagPGB Kp52145, ΔpagP::Km-GenBlock , RifR, KmR, the pagP gene inactivated
This study
52145-ΔpmrAB-ΔphoQGB 52145-ΔpmrAB, ΔphoQ::Km-GenBlock , RifR, KmR, the phoQ and pmrAB genes inactivated
This study
52145-ΔrcsB-ΔphoQGB 52145-ΔrcsB, ΔphoQ::Km-GenBlock , RifR, KmR, the phoQ and rcsB genes inactivated
This study
52145-ΔwcaK2-ΔpmrF 52145-ΔwcaK2, ΔpmrF , RifR, KmR, the pmrF gene inactivated in CPS mutant background
This study
52145-ΔwcaK2-Δ pagPGB 52145-ΔwcaK2, ΔpagP::Km-GenBlock , RifR, KmR, the pagP gene inactivated in CPS mutant background
This study
Plasmids pGEM-T Easy Cloning plasmid, AmpR (Promega) pGPL01 Firefly luciferase transcriptional fusion
suicide vector, AmpR (29)
pMAKSACB Suicide vector, Psc101 replication origin, Mob+, sacB gene, ClmR
(19)
pKOV Suicide vector, Psc101 replication origin, sacB gene, ClmR
(42)
pUC-4K Source GenBlock, AmpR, KmR (Pharmacia) pKD4 Km casette source for one-step
mutagenesis protocol. (14)
pGEMTFRTkm Km cassette source for mutagenesis This study
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 33 -
flanked by BamHI-FRT sites pFLP2 Plasmid encoding FLP to remove casettes
between FRT sites. Mobilizable. sacB for counterselection
(37)
pGEMTΔphoQGB pGEM-T Easy containing ΔphoQ::Km- GenBlock , AmpR, KmR
This study
pGEMTΔpagPGB pGEM-T Easy containing ΔpagP::Km- GenBlock , AmpR, KmR
This study
pGEMTΔrcsB pGEM-T Easy containing ΔrcsB, AmpR. This study pGEMTΔpmrABKm pGEM-T Easy containing ΔpmrABKm,
AmpR, KmR This study
pGEMTΔpmrD pGEM-T Easy containing ΔpmrD, AmpR This study pGEMTΔpmrDKm pGEM-T Easy containing ΔpmrDKm,
AmpR KmR This study
pGEMTΔpmrFKm pGEM-T Easy containing ΔpmrFKm, AmpR KmR
This study
pMAKSACBΔphoQGB pMAKSACB containing ΔphoQ::Km- GenBlock , ClmR, KmR
This study
pMAKSACBΔpagPGB pMAKSACB containing ΔpagP::Km- GenBlock , ClmR, KmR
This study
pMAKSACBΔrcsB pMAKSACB containing ΔrcsB, ClmR. This study pKOVΔpmrAB pKOV containing ΔpmrABKm, ClmR,
KmR This study
pKOVΔpmrD pKOV containing ΔpmrD, ClmR This study pKOVΔpmrF pKOV containing ΔpmrFKm, ClmR, KmR This study pGPLKpnPcps pGPL01 containing cps promoter region,
AmpR. This study
pGPLKpnPmrH pGPL01 containing pmrH promoter region, AmpR.
This study
pGPLKpnPagP pGPL01 containing pagP promoter region, AmpR.
This study
pGPLKpnMgtA pGPL01 containing mgtA promoter region, AmpR.
This study
pGPLKpnPmrD pGPL01 containing pmrD promoter region, AmpR.
This study
pGPLKpnPhoP pGPL01 containing phoP promoter region, AmpR.
This study
pGPLKpnRcsD pGPL01 containing rcsD promoter region, AmpR.
This study
pGPLKpnRcsC pGPL01 containing rcsC promoter region, AmpR.
This study
830
831
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 34 -
Table 2. Primers used in this study. 832 833
Sequences of primers used in this study Target gene Name Sequence (5’ to 3’) Mutagenesis phoP KpnphoPQmutr CCGGAATTCCGAACATCTCCCGGATATCG KpnphoPQmutf GTTCGATAAAGTCGGGCCAG rcsBF CGGGATCCCTGACGGTCCGCCGCCATGC rcsB rcsBR CGGGATCCCCAGCTGGAGAGCTGGGGAG rcsBinvF GACAACTGTCCATCCCCGATTC rcsBinvR TAATCAGCGTGATCCCGTCG KpnPagPF GACATTCACCATTACCGGGAAC pagP KpnPagPR TGCGCTGGTGGCTCGGCATG KpnPagPinvL CATTCCAGGTCTGCGCTACG KpnPagPinvR GGTTTCGGCGTCTCGCGCTG KpnpmrABR CAAGCTTGTGGCCAAAGCCATTGGCGAG pmrAB KpnpmrABF GGAATTCCAACGATAACGACGGCGGCTG KpnpmrABinvR GGGTACCCGCTACAGCCCTGAAGGGTCG KpnpmrABinvF GGGTACCCTCCCGCCGCATGCGGGAGAG MutpmrDupF TGGGCAAAGGTCGCCTGGTC pmrD MutpmrDupR CGGATCCGCGGCAGAGGCTGGCCGAAGC MutpmrDdownF CGGATCCGCTGAATAATAATCCGACGCAAAC MutpmrDdownR TCCAGCAAATAGTTGCGGAAC KpnpmrFF CGGATCCACCTGCGCGAGCTGGCGGAC pmrF KpnpmrFR CGGATCCCGGCGTCATCCGCGCCAATC KpnpmrFinvF TCTCCTCCGGCGGGTTTTGC KpnpmrFinvR CAAATACAGCTTTATGCGCCTG Promoter region cps K2ProcpsF GGAATTCCTGCTGGGACAAATTGCCACC K2ProcpsR AGATGGATGACCCCGCGATC phoP ProPhoPF GGGTACCCTCTTCATCCGGCAGGCCGAG ProPhoPR GGAATTCCGGCCGCCGAAGAAGGCTTCG rcsC ProrcsCF GGAATTCCATGAAGAGCTGGATGCCATCG ProrcsCR GGGTACCCGGGGTTCAAAGTTGGGCACC rcsD KpnPYojNF ATTTCCGCGCGCCGCGACTC KpnPYojNR GGAATTCCCGTTCGAGATCTCCGATTTGG pagP PKpnPagpF AGATAATGGCCGCGATGGAG KpnPagPinvR CATTCCAGGTCTGCGCTACG pmrD PropmrDF GGAATTCCTTTTATCTTCCTCCGGCAAAG PropmrDR TCTGAAGCACGACGCGGCAG pmrH PknppmrHF GGAATTCCGCGATGCCGGCCCGGCCTAC PkpnpmrHR GGGTACCGTCGCATGACGGTTGCCGGTC ugd PromkpnugdF GATCGTAGTCGGTCGGCGTG PromKpnugdR GGAATTCCAGTCCCAGAAAGGCGTATTGC mgtA ProKpnMgtAF GGAATTCCTCTTTGATGGTCAGCCGGTTC ProKpnMgtAR AGGTCCACAGATGCACCCACC Km cassette Cassette-F1 CGCGGATCCGTGTAGGCTGGAGCTGCTTCG Cassette-R1 CGCGGATCCCATGGGAATTAGCCATGGTCC RT-qPCR
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

- 35 -
pmrI KpnyfbGF1 CGCTGGATCTACTCGGTCTC KpnyfbGR1 TCTTTGTTCTCGATGATGCG cspE KpncspEF1 TCATTACTCCGGAAGATGGC KpncspER1 CCTTTGGCACCGTTAGTGAT rpoD KpnrpoDLEFT CCGGAAGACAAAATCCGTAA KpnrpoDRIGHT CGGGTAACGTCGAACTGTTT 834 835 836 837 838 839
on June 22, 2018 by guesthttp://iai.asm
.org/D
ownloaded from