a mi esposa a mis hermanos
TRANSCRIPT

1

Universidad Central “Marta Abreu” de Las Villas
Facultad de Ciencias Agropecuarias
UTILIZACIÓN DE STREPTOMYCES sp. RL8 COMO AGENTE
PROBIÓTICO EN POLLOS DE LA RAZA LEGHORN
Tesis presentada en opción al grado
científico de
Máster en Salud Animal Avanzada
Autor: Lic. Yoandry Martinez Arencibia
Tutores: Dr. C. Ricardo Medina Marrero
Dr. C. Milagro García Bernal
Santa Clara, Cuba 2018

Dedicatoria
A mi Mamá y mi Papá
A mi Esposa
A mis Hermanos

Agradecimientos
En pocas palabras quiero agradecer la colaboración de un grupo de personas sin
las cuales no hubiera sido posible culminar este trabajo.
En especial a (al):
Dr. C. Ricardo Medina Marrero y la Dr.C. Milagro R. García Bernal por
trasmitirme todas sus ideas, por su apoyo en el trabajo experimental y colaborar
en mi formación profesional.
MSc. Mirian Diaz Diaz y a la Téc. Marlen Casanova, por enseñarme los
primeros pasos en el mundo de la Microbiología, por su paciencia, guiarme y
trasmitirme sus conocimientos y experiencias.
René Dionicio Cupull, por su humildad, por su amplia capacidad formadora
de ideas, por darme sabios consejos, aclarar todas las dudas dándome su apoyo
incondicional y asesorarme en el trabajo experimental.
Lic. Eric Javier Prendes Rodríguez y Lic. Manuel Angel Soto Fexas, por ser
mis motivos de inspiración, por ser mis ojos, manos y mis pies, por guiarme a ser
mejor persona cada día. Gracias por ser mis amigos.
Dr.C. Leonel Lazo por brindarme sus consejos, ayuda desinteresada y
corregir mis errores.
Dr.C. Sergio Sifontes Rodríguez, por su paciente e importante ayuda en el
análisis de los resultados.
Téc. Igor Alvarez Herrera, por su amplia colaboración y conocimientos
hematológicos, asi como a la médico veterinaria Marisol Gutiérrez Parra.
Al personal del Área Biológica del Centro de Bioactivos Químicos,
especialmente a Jose, Yuly, Osmani, Zoe, Edisleidys, Meneses, Niurka, Cary y
Claudia, por su dedicación y colaboración, personas sin las que tampoco hubiera
sido posible lograr mis objetivos.
Molly, Yacel, Mileisy, Andresito y toda su familia por estar siempre
ayudándome en todo.
Mi familia (tías, tíos, primos, primas y parientes), amigos, colegas y a todas
las personas que me ayudaron y no han sido mencionadas.

Especialmente a:
Mi mamá y mi papá, gracias por elegirme a venir a este mundo, por su
gracia, humildad y amor, para que todo lo planeado se hiciese conforme a su
voluntad, gracias por demostrarme que sin ustedes no soy nadie, por ser mis
paradigmas como personas.
Diana, por ser lo mejor y más grande regalo que me ha dado la vida, por
ser mi otra hermana, amiga y compañera por convicción, por convertirse en mi
único y gran amor.
A mi tio Juan, Ernesto, Yake, Luz Clara, La Poty y Favy.
Rome, Gleivys, Anibal, Zaida, Tania, Amanda, Isaac, Mary Luz y Luz
Marina por acogerme como otro miembro más de la familia.
A mis hermanos por quererme siempre sin importar nada ni nadie, por
darme ese amor incondicional que nos caracteriza. A Ita, Pingui, Mily y Yuny, que
son mis grandes amores.

Índice
INTRODUCCIÓN………………………………………………………………………. 1
CAPÍTULO 1. REVISIÓN BIBLIOGRÁFICA………………………………………. 4
1.1 Producción y consumo de la carne de pollo en el mundo……………… 5
1.2 Enfermedades en la avicultura…...……………………………..………... 6
1.2.1 Escherichia coli……………………………………………………..………. 7
1.2.2 Colibacilosis………………………………………………………..……….. 7
1.3 Antibióticos utilizados en la avicultura………………………….…………. 9
1.3.1 Resistencia a antibióticos………………………………………………… 9
1.4 Probióticos…………………………………………………………………. 11
1.4.1 Característica de un microorganismo para candidato a probiótico… 11
1.4.1.1 Antagonismo frente a diferentes patógenos…………………………… 11
1.4.1.2 Resistencia al pH y a las bilis……………………………………………… 11
1.4.1.3 Inocuidad…………………………………………………………………….. 11
1.4.1.4 Capacidad de adhesión a células intestinales…………………………… 11
1.4.2 Mecanismos de acción de los probióticos……………………………….. 12
1.4.2.1 Producción de compuestos inhibitorios………………………………….. 12
1.4.2.2 Competencia por sitios de adhesión o exclusión competitiva…………. 12
1.4.2.3 Competencia por nutrientes………………………………………………... 12
1.4.2.4 Modulación del sistema inmunológico………………………………........ 13
1.4.2.5 Mejoras en la calidad del agua…………………………………………….. 13
1.4.3 Uso de Probióticos en la avicultura……………………………………….. 13
1.5 Características generales de los actinomicetos como candidatos a
probióticos…………………………………………………………………….. 14
1.5.1 Actinomicetos utilizados como promotores del crecimiento en la
producción avícola…………………………………………………………… 15

CAPÍTULO 2. ESTUDIO DE LA TOLERANCIA DE LA CEPA
STREPTOMYCES sp. RL8 A SALES BILIARES, CLORURO DE SODIO Y pH. 19
2.1 Materiales y Métodos……………………………………………………… 20
2.1.1 Determinación de la tolerancia a diferentes valores de pH………........ 21
2.1.2 Determinación de la tolerancia a diferentes concentraciones de sales
biliares ……………………………………………………………………… 21
2.1.3 Determinación de la tolerancia a diferentes concentraciones de cloruro
de sodio………………………………………………………………………. 21
2.2 Resultados y Discusión………………………………………….……........ 21
2.2.1 Determinación de la tolerancia a diferentes valores de pH……………. 21
2.2.2 Determinación de la tolerancia a diferentes concentraciones de sales
biliares …………………………………………….………………………… 24
2.2.3 Determinación de la tolerancia a diferentes concentraciones de cloruro
de sodio ……………………………………………………………………… 25
2.3 Conclusiones parcilaes…………………………………………………….. 26
CAPÍTULO 3. EVALUACIÓN DEL EFECTO DE LA CEPA STREPTOMYCES
sp. RL8 SOBRE LOS PARÁMETROS tBIOPRODUCTIVOS Y DE SALUD…… 27
3.1 Materiales y Métodos………………………………………………………. 28
3.2 Resultados y Discusión……………………………………………………. 30
3.3 Conclusiones parciales…………………………………………………….. 33
CAPÍTULO 4. ESTUDIO DE LOS INDICADORES HEMATOLÓGICOS AL
EVALUAR LA CEPA DE STREPTOMYCES sp. RL8 Y SU POSIBLE EFECTO
MODULADOR DEL SISTEMA INMUNE EN POLLOS RAZA LEGHORN……………… 34
4.1 Materiales y Métodos……………………………………………………….. 35
4.2 Resultados y Discución…………………………………………………….. 36
4.3 Concluciones parciales……………………………………………………… 38
CAPÍTULO 5. DETERMINACIÓN DE LAS TRANSFORMACIONES
MICROBIOLÓGICAS EN EL TRACTO DIGESTIVO DE LAS AVES EVALUADAS……. 40
5.1 Materiales y Métodos………………………………………………………... 41

5.2 Resultados y Discusión……………………………………………………. 41
5.3 Conclusiones parciales…………………………………………………….. 43
CONCLUSIONES GENERALES…………………………………………………….. 44
RECOMENDACIONES……………………………………………………………….. 46
REFERENCIAS BIBLIOGRÁFICAS………………………………………………… 48
ANEXOS………………………………………………………………………………… 59

Resumen
En la producción animal, la introducción de probióticos en los sistemas de
alimentación y manejo constituye una alternativa para incrementar el estado de
salud de los animales, disminuir el uso indiscriminado de antibióticos y
consecuentemente, la eficiencia productiva y económica. El presente trabajo tuvo
como objetivo evaluar la actividad probiótica de Streptomyces sp. RL8 en
indicadores fisiológicos y bioproductivos en pollos Leghorn. Estos
microorganismos se caracterizaron in vitro según su capacidad de crecimiento a
diferentes valores de pH ácidos, así como su tolerancia a altas concentraciones
de sales biliares y a cloruro de sodio. La inclusión in vivo de este microorganismo
en la dieta de los pollos produjo una respuesta de tipo probiótica en indicadores
de salud y crecimiento de las aves, se encontraron diferencias significativas entre
ambos grupos para algunos componentes hematológicos e inmunológicos. Se
produjo una disminución significativa de los colibacilos en el tracto gastrointestinal
(TGI), además de una buena colonización intestinal de la cepa utilizada. Los
resultados obtenidos in vitro e in vivo, demuestran que la cepa Streptomyces sp.
RL8 posee propiedades probióticas y puede ser utilizada como aditivo microbiano
destinado a la alimentación de pollos recién eclosionados, para controlar las
poblaciones de colibacilos hasta los 21 días de edad en pollos de remplazo de la
raza Leghorn.
Palabras claves: Streptomyces sp. RL8, probiótico, colibacilos

Abstract
In animal production, the introduction of probiotics in feeding and management
systems constitutes an alternative to increase the health status of animals,
decrease the indiscriminate use of antibiotics and, consequently, bioproductive
and economic efficiency. The objective of this work was to evaluate the probiotic
activity of Streptomyces sp. RL8 in physiological and bioproductive indicators in
Leghorn chickens. These microorganisms were characterized in vitro according to
their growth capacity at different acidic pH values, as well as their tolerance to high
concentrations of bile salts and sodium chloride. The in vivo inclusion of this
microorganism in the diet of the chickens produced a probiotic response in
indicators of health and growth of the birds, significant differences were found
between both groups for some hematological and immunological components.
There was a significant decrease of the colibacilos in the gastrointestinal tract
(GIT), in addition to a good intestinal colonization of the strain used. The results
obtained in vitro and in vivo show that the strain Streptomyces sp. RL8 has
probiotic properties and can be used as microbial additives used to feed newly
hatched chickens, to control colibacillus populations up to 21 days of age in
replacement chickens of the Leghorn breed.
Key words: Streptomyces sp. RL8, probiotic, colibacilos

1
INTRODUCCIÓN

2
INTRODUCCIÓN
En las instalaciones avícolas los pollitos eclosionan en un medio casi estéril y son
ubicados en sus primeras semanas de vida en galpones con condiciones
higiénicas estables, que imposibilitan la colonización de una microbiota intestinal
beneficiosa; además de esto, los pollos están sometidos a situaciones que les
producen estrés como son: la alta densidad de población, vacunación, altas o
bajas temperaturas, humedad inadecuada, incidencia de gases tóxicos,
inmunodepresión; que ocasionan desbalances de la microbiota intestinal en caso
de que esta exista. De esta manera las aves se encuentran expuestas a la
incidencia de microorganismos patógenos que pueden causar enfermedades
gastrointestinales como la salmonelosis y la colibacilosis. Para contrarrestar los
efectos de dichas enfermedades se utilizan los antibióticos de amplio espectro;
pero estos compuestos han originado graves problemas de resistencia microbiana
y efectos residuales colocando en riesgo la inocuidad alimentaria. Por otro lado,
se aumentan los costos del modelo de producción y una notable disminución en
los índices bioproductivos de la industria avícola (Chichlowski et al., 2007).
Por lo anteriormente expuesto se plantea la utilización de probióticos como
coadyuvantes dietéticos de origen microbiano que benefician la fisiología del
hospedante al modular la inmunidad de la mucosa y la inmunidad sistémica, así
como mejorar el balance nutricional y microbiano en el TGI; en la producción
avícola, se reportan resultados sobre el uso de estos aditivos microbianos que
como en la mayoría de los casos, suelen ser muy satisfactorios por los buenos
rendimientos en la producción (Ongor et al., 2008). Entre los productos probióticos
con propósito aviar, se destacan aquellos que contienen microorganismos
autóctonos del TGI de aves en estado saludable constituidas principalmente por
diferentes especies de los géneros bacterianos: Lactobacillus, Bacillus,
Estreptococos, Enterococos y levaduras Saccharomyces; los cuales han
demostrado una alta eficiencia en la reducción de patógenos intestinales y en el
aumento de los indicadores bioproductivos (Byrd et al., 2003).
En Cuba, no se emplean estos productos, pero existe la experiencia
biotecnológica necesaria para desarrollarlos con tecnologías económicamente
viables, y que a su vez, mejoren el rendimiento bioproductivo y la salud de los
animales (Collins & Gibson, 1999; Pérez et al., 2011). En nuestro país no se han
realizado estudios donde se utilice como aditivo probiótico los estreptomicetos

3
como agentes controladores de la colibacilosis y otras enfermedades zoonóticas
de origen bacteriano, cuando se conoce que los actinomicetos representan una
fuente de metabolitos bioactivos para combatir la resistencia bacteriana de
antibióticos usados comúnmente (Lino-Navarro et al., 2016).
Problema científico
Se desconoce el efecto de la utilización de Streptomyces sp. RL8 como agente
probiótico en pollos de 21 días de edad de la raza Leghorn.
Hipótesis
Es posible controlar la colonización por E. coli a nivel de intestino delgado y
grueso en pollos de 21 días de edad, mediante la utilización de Streptomyces sp.
RL8 como agente probiótico en el remplazo de gallinas ponedoras raza Leghorn.
Objetivo general
Evaluar la actividad probiótica de Streptomyces sp. RL8 en indicadores
fisiológicos y bioproductivos en pollos de remplazo de la raza Leghorn.
Objetivos específicos
1. Determinar in vitro la tolerancia a pH, sales biliares y cloruro de sodio de la
cepa Streptomyces sp RL8.
2. Evaluar in vivo el efecto de la cepa de Streptomyces sp. RL8 sobre los
parámetros bioproductivos, de salud y morfométricos sobre en las aves
evaluadas.
3. Caracterizar el efecto de la cepa Streptomyces sp. RL8 en los parámetros
hematológicos y su posible efecto modulador sobre el sistema inmune de las aves
evaluadas.
4. Determinar las transformaciones microbiológicas en el tracto digestivo de
las aves evaluadas.

4
CAPÍTULO 1.
REVISIÓN
BIBLIOGRÁFICA

5
CAPÍTULO 1. REVISIÓN BIBLIOGRÁFICA
1.1 Producción y consumo de la carne de pollo en el mundo
De acuerdo con el reporte Perspectivas Agrícolas 2016‐2025 de la Organización
para la Cooperación y el Desarrollo Económico (OCDE) y la Organización de las
Naciones Unidas para la Alimentación y la Agricultura (FAO/WHO), se proyecta
que la carne de ave encabece el crecimiento en la producción mundial de carne,
debido al incremento en la demanda mundial de esta proteína animal, en
comparación con las carnes rojas. Así, se estima que el consumo de carne de ave
represente dos tercios del consumo adicional del total de carnes. Los bajos costos
de producción y un accesible precio del producto, en comparación con otros tipos
de carne, han contribuido a que la carne de ave se convierta en una importante
opción de consumo en los países en desarrollo. Por otra parte, la producción
intensiva, la alta conversión alimenticia y el ciclo de producción más corto de las
aves, permiten que la avicultura sea la actividad que más rápido incorpore los
beneficios de la reducción de los precios de los granos. Así, entre 2016 y 2025, la
producción y el consumo crecerán a una tasa promedio anual de 1,5 porciento,
ubicándose en ambos casos en 131,2 millones de toneladas en 2025
(USDA‐FAS, 2016).
La producción mundial de pollo creció un 2 % en 2018 a 92,5 millones de
toneladas, principalmente a partir de las ganancias en los Estados Unidos, Brasil,
India y la Unión Europea (UE). La expansión de los Estados Unidos y Brasil se ve
reforzada por la abundancia de suministros de alimentos a precios relativamente
bajos, la ausencia de influenza aviar altamente patógena (HPAI) y la modesta
demanda mundial. Las ganancias en la Unión Europea y la India se deben a la
creciente demanda interna. Luego de dos años de declive, la producción de China
ahora se pronostica se incremente levemente (un 1 %) la producción de pollos
debido a los esfuerzos para frenar los brotes de influenza aviar altamente
patógena (IAAP), los costos de alimentación estables y el rebote de la demanda
interna. Sin embargo, la expansión sigue parcialmente limitada por la falta de
nuevas reservas genéticas debido a prohibiciones relacionadas con la IAAP en la
mayoría de los principales proveedores (USDA‐FAS, 2018).
Las exportaciones mundiales en 2018 se pronostican un 2 % más altas con un
récord de 11,3 millones de toneladas, respaldadas por una mayor demanda de

6
una diversidad de mercados, en particular Japón, Cuba, Hong Kong, Angola, Irak
y Ghana. En Brasil se pronostica un 1 % más alto con un récord de 3,9 millones
de toneladas, ya que los desafíos de acceso al mercado en Arabia Saudita y
China serán más que compensados por los mayores envíos a otros mercados. Se
pronostica que Tailandia tendrá un aumento del 7 % hasta 810,000 toneladas
debido a la fuerte demanda en Asia y un mayor acceso a los mercados de China.
Los envíos de Ucrania aumentarán un 18 % hasta 310,000 toneladas debido a la
demanda en el Medio Oriente, la Unión Europea y el Norte de África (USDA‐FAS,
2018).
Los principales países exportadores de pollo son Brasil, Estados Unidos, Unión
Europea, Tailandia, y China. En conjunto estos cinco estados concentran el 86,8
% de las exportaciones mundiales de carne de pollo, siendo Brasil y Estados
Unidos los líderes mundiales con una participación de dos tercios del total
(USDA‐FAS, 2016).
1. 2 Enfermedades en la avicultura
La contaminación microbiana de la carne de pollo es indeseable pero inevitable, y
depende de la calidad microbiológica de las canales utilizadas como materia
prima. Las prácticas de higiene durante la manipulación, el tiempo y la
temperatura de almacenamiento afectan de forma importante el crecimiento
microbiano. En la carne de aves se han encontrado varios cientos de especies de
microorganismos. Estos pueden dividirse en dos grupos generales, por una parte,
los que son capaces de producir enfermedades en humanos, generalmente
denominados patógenos, y por otra la alteración de la carne, conocidos como
microorganismos alterantes. Una de las principales fuentes de Salmonella y E.
coli en la carne de los pollos lo constituyen los piensos y harinas conjuntamente
con sus subproductos de origen animal y el agua. Los insectos pueden ser
reservorios y vectores de microorganismos. Los pelos, heces de roedores y otros
pequeños mamíferos, hacen que los microorganismos se propaguen dentro de los
corrales avícolas. Las aves silvestres o aves domésticas también pueden
transmitir Salmonella, E. coli y otros microorganismos de granja a granja (Pérez,
2015).

7
1.2.1 Escherichia coli
E. coli pertenece a la familia de las enterobacterias. Las cepas productoras de
enfermedad diarreica, se clasifican en distintos grupos por sus características
patogénicas: Se han descrito seis patotipos de E. coli involucrados en procesos
diarreicos, mediante la identificación de factores de virulencia y mecanismos de
patogenicidad, E. coli enteropatógena, E. coli enterotoxigénica (ECET), E. coli
enteroinvasora (ECEI), E. coli shigatoxigénica (ECST), E. coli enteroagregativa
(ECEA), E. coli adherente difusa (ECAD) y E coli adherente invasora (ECAI)
(Farfán-García et al., 2016). Esta bacteria es habitante habitual del intestino de
todos los animales. La supervivencia en los alimentos oscila de diez días a dos
meses, incluso a pH bajos o en congelación. Poco tolerante al calor (se inactiva a
68,3ºC/40seg) (Pérez, 2015).
La E. coli es un habitante común del intestino de las aves de corral, puede
encontrarse 109 unidades formadoras de colonias de bacterias por gramo (UFC/g)
de heces; también se ha aislado E. coli del tracto respiratorio, piel y plumas tanto
de aves enfermas como sanas. En la microbiota cecal de pollos sanos de diez a
15 % de las cepas de E. coli son potencialmente patógenas (Rocha, 2012).
1.2.2 Colibacilosis
Se refiere a cualquier infección localizada o sistémica causada total o
parcialmente por E. coli patógena aviar, que incluye colisepticemia, septicemia
hemorrágica, coligranuloma (enfermedad de Hjarre), enfermedad del saco aéreo
(enfermedad respiratoria crónica, CRD), síndrome de la cabeza hinchada,
venéreo colibacilosis, celulitis coliforme (procesos inflamatorios o infecciosos),
peritonitis, salpingitis, orquitis, osteomielitis, sinovitis (incluido el complejo de
osteomielitis de pavo), panoftalmitis, infección por omalisto, saco vitelino y
enteritis. A menudo, la colibacilosis es una de las enfermedades más frecuentes
en las encuestas sobre la salud de las aves de corral o en las cadenas de
procesamiento. Otros agentes infecciosos y factores no infecciosos generalmente
predisponen a un ave a la infección o contribuyen a la gravedad de la
enfermedad. Dos especies adicionales de Escherichia, E. fergusonii y E. albertii,
han sido aisladas de aves y son capaces de causar enfermedad o son de
importancia para la salud pública. E. coli no posee capacidades de resistencia
únicas y tiene un patrón de susceptibilidad a los agentes químicos y físicos típicos
de bacterias vegetativas, Gram negativas (Gornatti-Churria et al., 2018).

8
Esta enfermedad infecciosa, producida por la E. coli, es considerada una de las
principales causas de morbilidad en la avicultura, e incluso, llegando a estar por
encima del 20 % de mortalidad. Las enfermedades generadas por E. coli son
responsables de la mayoría de las pérdidas económicas por causa bacteriana
para la industria aviar en el mundo. Aunque existen más de 170 serotipos de E.
coli, los más comúnmente encontrados produciendo enfermedad en las aves son
el O1, O2 y O78; la identificación serológica es además de complicada costosa y
por lo tanto es difícil determinar con exactitud el serotipo. La colibacilosis incluye
una variedad de infecciones localizadas o sistémicas, donde E. coli puede estar
involucrada como el agente infeccioso primario o secundario. La infección con E.
coli en mamíferos frecuentemente afecta el intestino, sin embargo, en aves, las
infecciones más notables ocurren fuera de este órgano. Tanto pollos de engorde,
como ponedoras pueden sufrir de salpingitis aguda o crónica, la cual puede
resultar de una infección de tipo ascendente desde la cloaca, o de una infección
del saco aéreo abdominal izquierdo, esta salpingitis puede llevar a una pérdida de
la capacidad de postura en las gallinas ponedoras. Una gallina ponedora que
padezca de ooforitis o salpingitis inducida por E. coli, puede infectar sus huevos
antes de la formación de la cáscara, a su vez existe riesgo de contaminación fecal
durante el paso del huevo a través de la cloaca y también luego de ser puesto;
esta última posibilidad es considerada como la principal ruta de infección para el
huevo. Antes de la incubación, las E. coli patogénicas causan infecciones del
saco vitelino y mortalidad embrionaria. En pollos de engorde la E. coli patogénica
puede producir dermatitis necrótica, también conocida como celulitis, que se
caracteriza por una inflamación crónica del tejido celular subcutáneo en el
abdomen y los muslos (Rocha, 2012).
Cuando la E. coli alcanza el sistema vascular, infecta los órganos internos y el
corazón, pudiendo causar falla cardiaca por afección del miocardio.
Ocasionalmente también lleva a sinovitis y osteomielitis y en raras ocasiones a
panoftalmia. El coligranuloma o enfermedad de Hjarre se caracteriza por la
presencia de granulomas en el hígado, los ciegos, duodeno y mesenterio. Esta es
una rara forma de colibacilosis, pero en granjas afectadas puede causar hasta un
75 % de mortalidad en aves (Rocha, 2012).

9
1.3 Antibióticos en la avicultura
Durante décadas, los antibióticos se utilizaron en la producción animal como
aditivos promotores del crecimiento para incrementar los rendimientos
bioproductivos. Sin embargo, su uso creó serios problemas de resistencia
microbiana y agudizó la aparición de efectos residuales en los alimentos para el
consumo humano. Por tales razones, fue necesario la introducción en las
prácticas de alimentación y manejo de otros aditivos como los probióticos que
contribuyeran a evitar los efectos negativos del uso de antibióticos (García, 2011).
1.3.1 Resistencia a antibióticos
En las aves de corral, los antibióticos y los antiparasitarios se usan ampliamente
para la prevención y el tratamiento de enfermedades. En los Estados Unidos, los
antibióticos se utilizan para la promoción del crecimiento, aunque este tipo de uso
se ha prohibido en la Unión Europea desde 2006, ya que estos pueden contener
residuos de medicamentos veterinarios y presentar riesgos para la salud humana,
incluidos efectos tóxicos directos, reacciones alérgicas y una mayor resistencia
bacteriana a antibióticos comunes (Goetting et al., 2011).
La resistencia a los antimicrobianos es un fenómeno biológico natural. Cada vez
que se ha puesto en uso un nuevo agente antimicrobiano en el ámbito clínico. La
resistencia se entiende como el mecanismo mediante el cual la bacteria puede
disminuir la acción de los agentes antimicrobianos. Desde el punto de vista
clínico, se considera que una bacteria es sensible a un antibacteriano cuando la
concentración de éste en el lugar de la infección es al menos cuatro veces
superior a la Concentración Inhibitoria Mínima (CIM). Una concentración por
debajo de la CIM clasifica a las bacteria como resistentes y valores intermedios
como moderadamente sensibles (Rocha, 2012).
La resistencia a los antimicrobianos afecta a la gran mayoría de los géneros y
especies microbianas que pueden ser transferidos al hombre desde el ambiente
por diferentes vías, entre ellas, la cadena alimentaria. Salmonella y E. coli, que
con frecuencia afectan al hombre y también a los animales, se consideran los
géneros con mayores riesgos de transferencia zoonótica de resistencias. La
contaminación de los alimentos durante la manipulación, puede ser directa entre
los alimentos crudos o indirecta a través de las manos, superficies y utensilios
contaminados. El TGI de los animales y humanos es un ecosistema favorable

10
para el intercambio de los genes de resistencia entre géneros e interespecies
(Rocha, 2012).
En Cuba y el mundo, se informan para Salmonella y E. coli aislados de alimentos,
altos porcentajes de resistencia a antibióticos de uso común como la tetraciclina y
la ampicilina. La resistencia bacteriana se produce por una combinación de
factores entre los que se destaca el uso inadecuado de los antimicrobianos en el
hombre y en los animales. En los animales, estos fármacos se utilizan con tres
finalidades: terapéutica, profiláctica y como promotores del crecimiento. La
utilización de los antibióticos como promotores del crecimiento animal constituye
una práctica conocida desde la década de 1950, momento en el que se descubre
que pequeñas dosis de tetraciclina mejoraban el desarrollo. En aquel momento,
se desestimó el efecto que el consumo de estos “factores nutricionales” pudiera
tener sobre la resistencia; de esta forma, fármacos como la tetraciclina, la
penicilina y el cloranfenicol, medicamentos destinados al tratamiento de
infecciones humanas comienzan también a utilizarse con fines de engorde animal,
y no es hasta los inicios de la década de los años 70 que surgen las primeras
alertas relacionadas con esta práctica, al observarse en Salmonella un incremento
de la resistencia al cloranfenicol (Puig et al., 2011).
Se describe el incremento en la resistencia a las quinolonas en patógenos como
E. coli y Campylobacter spp., tanto de origen animal como humano, efecto que se
atribuye al amplio consumo de dichos fármacos en la profilaxis y la terapéutica
animal (Puig et al., 2011).
Los residuos de medicamentos en huevos de gallina son motivo de preocupación
porque relativamente pocas drogas, están etiquetadas para gallinas ponedoras,
aunque varios medicamentos están aprobados para otras clases de producción
de aves de corral (Castanon, 2007). Los residuos de medicamentos en los huevos
pueden surgir cuando las gallinas ponedoras reciben por error alimento medicado,
cuando el alimento se contamina en el molino durante la mezcla, o cuando los
medicamentos se dan fuera de etiqueta (Kennedy et al., 2000). Mientras que un
pollo pone un huevo aproximadamente cada 24 h, cada huevo tarda varios días
en desarrollarse in vivo, y algunos componentes del huevo existen meses antes
de que el huevo este completamente desarrollado y sin cáscara que los contiene
se coloque (Etches, 1996; Whittow, 2000). Debido a la naturaleza prolongada del
desarrollo del huevo, pueden requerirse muchas semanas después del

11
tratamiento o la exposición antes de que los huevos estén libres de residuos de
medicamentos (Goetting et al., 2011).
1.4 Probióticos
Según Mehdi et al,. (2018) los probióticos se definen como microorganismos que,
cuando se administran en cantidades adecuadas, confieren un beneficio para la
salud en el huésped.
1.4.1 Característica de un microorganismo para candidato a probiótico
De forma general las principales características se resumen a alta hidrofobicidad
de la superficie celular, coagregación y resistencia a las sales biliares y también
condiciones ácidas (Taheri et al., 2009).
1.4.1.1 Antagonismo frente a diferentes patógenos
Esta es una vía muy común para la primera selección de posibles probióticos. Las
cepas en estudio son expuestas in vitro a los patógenos seleccionados, o a sus
productos extracelulares. Dicho efecto puede evaluarse en medio líquido o sólido
(Irianto & Austin, 2002).
1.4.1.2 Resistencia al pH y a la bilis
Dentro de las pruebas de selección in vitro se incluye la resistencia a la acidez
gástrica y a las sales biliares, que constituyen condiciones limitantes para la
supervivencia a través del TGI, de lo contrario los microorganismos probióticos no
llegarían viables al final del intestino para ejercer su acción beneficiosa para la
salud del hospedero (Kesarcodi-Watson et al., 2008).
1.4.1.3 Inocuidad
Una cepa probiótica debe estar exenta de patogenicidad y ser incapaz de
ocasionar daño al hospedero, además de ofrecer efectos beneficiosos mediante
su interacción con el mismo (Schrezenmeir & de Vrese, 2001). Al mismo tiempo,
debe ser reconocida como GRAS (generalmente considerada como segura, por
sus siglas en inglés) (FAO/WHO, 2002).
1.4.1.4 Capacidad de adhesión a células intestinales
Para sobrevivir y competir en un ecosistema complejo como el intestino, es
necesario que la cepa seleccionada pueda adherirse a la mucosa intestinal del
hospedero, siendo este un criterio importante a la hora de evaluar una cepa como
probiótica. Solo las cepas capaces de adherirse podrían llegar a colonizar el
intestino, y de esa manera formar parte de una primera barrera defensiva frente a

12
los patógenos. En caso contrario la cepa se convertirá en un organismo
transitorio, limitando sus posibles efectos positivos y haciendo indispensable su
ingestión periódica (Vine et al., 2006).
1.4.2 Mecanismos de acción de los probióticos
Los probióticos generalmente modulan el crecimiento de la microbiota intestinal,
suprimiendo a las bacterias potencialmente perjudiciales y reforzando los
mecanismos de defensa naturales del hospedero (Giraffa et al., 2010), mejorando
así su resistencia contra las enfermedades infecciosas (Gildberg et al., 1997).
1.4.2.1 Producción de compuestos inhibitorios
Los probióticos juegan un papel importante en la prevención de la aparición de
enfermedades mediante la producción de ciertos compuestos inhibitorios que
actúan antagónicamente contra los microorganismos patógenos y por lo tanto,
evitan su proliferación en el hospedero (Tinh et al., 2007). La actividad de un
probiótico contra diversos agentes patógenos puede ser debida a la producción
de antibióticos, bacteriocinas, sideróforos, lisozimas, proteasas y peróxido de
hidrógeno (Bernal et al., 2015).
1.4.2.2 Competencia por sitios de adhesión o exclusión competitiva
La competencia por sitios de adhesión sobre el mucus del TGI y otras superficies
de tejidos, es un posible mecanismo para prevenir su colonización por patógenos.
La adhesión bacteriana a la superficie de un tejido determinado, es una etapa
importante para la infección patógena, por lo tanto para que un probiótico ejerza
su acción benéfica, debe competir con los patógenos por los sitios de adhesión,
ejerciendo así un efecto de exclusión competitiva (Verschuere et al., 2000).
La exclusión competitiva consiste en la prevención o reducción de la colonización
de una bacteria patógena, por parte de las bacterias presentes en el TGI del
hospedero. Las bacterias probióticas se establecen en los sitios de adhesión de la
mucosa intestinal, formando una barrera física y de esa manera limitan o evitan el
establecimiento de las bacterias patógenas en el intestino (Lara-Flores, 2011).
1.4.2.3 Competencia por nutrientes
Una vez suministrados, los probióticos más competentes constituyen parte de la
microbiota residente del TGI del hospedero. Lo cual se logra mediante su
adhesión duradera o permanente a la mucosa, a las células epiteliales y a otros
tejidos, para mantener e incrementar la salud o el bienestar del hospedero

13
(Gatesoupe, 1999). La capacidad de adhesión de algunas bacterias se probó in
vitro e in vivo y sus resultados sugieren que el patógeno es desplazado por el
potencial probiótico como resultado de la competencia por espacio y nutrientes
esenciales (Verschuere et al., 2000).
1.4.2.4 Modulación del sistema inmunológico
Las bacterias probióticas tienen una acción estimulante sobre el sistema
inmunitario del hospedero, ya que actúan sobre las células implicadas tanto en la
inmunidad natural como en la inmunidad específica. Estos microorganismos
activan a los macrófagos, favorecen la producción de anticuerpos y estimulan la
respuesta defensiva ante virus, bacterias y parásitos que provocan enfermedades.
Muchos probióticos han sido identificados por su acción inmunomoduladora, al
mejorar la actividad fagocítica, el estallido respiratorio y la actividad superóxido
dismutasa y peroxidasa, además de incrementar el número de leucocitos,
linfocitos y eritrocitos en vertebrados (Newaj-Fyzul et al., 2014).
1.4.2.5 Mejoras en la calidad del agua
Las bacterias Gram positivas, tales como Bacillus spp., estimulan el sistema
inmune del hospedero y también actúan mejorando la calidad del agua. Bacillus
spp., actúa de manera más eficiente en la conversión de materia orgánica en
dióxido de carbono, mientras que las bacterias Gram negativas convierten mayor
proporción de materia orgánica en biomasa bacteriana o en limo (Nemutanzhela
et al., 2014).
1.4.3 Uso de Probióticos en la Avicultura
Existen diferentes investigaciones donde los utilizan diversos microorganismos
como agentes probióticos en el control de enfermedades. Las bacterias ácido
lácticas se encuentran entre los microorganismos que más se estudian y utilizan
como candidatos probióticos o en productos probióticos, fundamentalmente los
del género Lactobacillus. También se emplean Streptococcus, Enterococcus,
Pediococcus, Leuconostoc, Bacillus, Bifidobacterium, Propionibacterium,
Bacteroides, E. coli (cepas no patógenas), Aspergillus y levaduras (García, 2011).
La eficacia de los preparados probióticos se puede incrementar cuando se
seleccionan cepas más eficientes; se emplean mezclas de cepas; se realizan
manipulaciones genéticas o cuando se combinan probióticos y componentes
sinérgicos como los prebióticos (Kazue et al., 2015).

14
1.5 Características generales de los actinomicetos como candidatos a
probióticos
Los actinomicetos son microorganismos abundantes en el suelo, sin embargo, se
encuentran también en ambientes acuáticos, tanto dulces como marinos. Entre
sus características particulares se encuentra la de producir un olor típico a suelo
húmedo, debido a la producción de un metabolito denominado geosmina.
Además, se caracterizan por presentar una actividad metabólica alta, y producir
terpenoides, pigmentos y enzimas extracelulares que les confieren una gran
capacidad para degradar la materia orgánica de origen vegetal y animal. Los
actinomicetos constituyen un grupo heterogéneo de microorganismos; son
bacterias Gram positivas que se caracterizan por su capacidad de formar
filamentos ramificados. El orden de los Actinomycetales comprende 63 géneros,
constituyendo aproximadamente del 20 al 60 % de la población microbiana total
del suelo (Bernal, 2016).
Los metabolitos secundarios obtenidos de Actinomycetales proporcionan una
fuente potencial de muchos compuestos nuevos con propiedades antibacterianas,
antitumorales, antifúngicas, antivirales, antiparasitarias y de otro tipo. La mayoría
de estos compuestos se usan ampliamente como medicamentos para combatir
cepas bacterianas Gram positivas y Gram negativas resistentes a múltiples
fármacos. Los miembros del género Streptomyces spp., son productores de
metabolitos secundarios bioactivos (Solecka et al., 2012).
Actualmente, los actinomicetos se encuentran incluidos en el dominio bacteria
debido a las siguientes razones: su pared celular está compuesta por
péptidoglicanos, el diámetro de sus hifas (0,5 a 2, μm), son sensibles a los
antimicrobianos, pero presentan resistencia a los antifúngicos y la disposición de
su material genético es típicamente procariótico (Procópio, 2012). Se caracterizan
por presentar un alto contenido de guanina y citosina en su ADN, encontrándose
entre el rango de 51 a 78 % G+C. Estas bacterias son generalmente aerobias,
pero algunas son anaerobias, pudiéndose encontrar en animales o en el hombre;
son heterótrofas, por lo cual pueden utilizar fuentes de carbono simples o
complejas, y compuestos moleculares orgánicos tales como ácidos, azúcares,
polisacáridos, lípidos, proteínas e hidrocarburos alifáticos. Los actinomicetos
utilizan como fuentes de nitrógeno amonio, nitratos, aminoácidos, peptonas y un
gran número de proteínas (Bernal, 2016).

15
Los actinomicetos son los miembros más importantes en el mundo microbiano por
su potencial para producir compuestos biológicamente activos. Alrededor del 45
% de todos los compuestos bioactivos obtenidos a partir de microorganismos son
producidos por actinomicetos (Bernal, 2016) .Los antibióticos utilizados en la
práctica clínica tales como la fosfomicina, lincomicina, neomicina, estreptomicina,
daptomicina, la eritromicina y la tetraciclina son producidos por el género
Streptomyces perteneciente al Orden Actinomycetales (Gupta et al., 2015).
Streptomyces es un género de bacterias Gram positivas, que crece en diferentes
entornos y su forma se asemeja a los hongos filamentosos. La diferencia
morfológica de Streptomyces implica la formación de una capa de hifas que
pueden diferenciarse en una cadena de esporas. Los miembros de este género
son habitualmente reconocidos como microorganismos GRAS (según sus siglas
en inglés) (Lannot, 2005).
1.5.1 Actinomicetos utilizados como promotores del crecimiento en la
producción avícola
En la década de 1940, el uso de probióticos Streptomyces aureofaciens provocó
un aumento del peso en los animales, lo que llevó al descubrimiento de la
clortetraciclina. Las tetraciclinas, macrólidos, avoparcina y penicilinas se han
usado comúnmente en la ganadería para promover el crecimiento a través del
aumento de la ingesta de alimentos, el aumento de peso y la mejora de la salud
(Angelakis, 2017).
Tanaka et al., (1975) descubrieron las nanaomicinas, antibióticos nuevos
producidos por la cepa OS-3966 que se designó Streptomyces rosa var.
notoensis. Las propiedades físicas y químicas sugieren que las nanaomicinas A y
B son compuestos relacionados con la quinona. Las nanaomicinas A y B inhiben
principalmente micoplasmas (Mycoplasma gallisepticum es el agente patógeno
más importante, desde el punto de vista económico, en la producción avícola),
hongos y bacterias Gram positivas.
Danforth et al., (1977) trataron infecciones con especies únicas de Eimeria
acervulina, E. mivati, E. maxima, E. tenella, E. necatrix y E. brunetti, donde
realizaron en tres experimentos por separados para evaluar la eficacia
anticoccidiana de varios niveles de salinomicina (AHR-3096), un producto de
fermentación de Streptomyces albus. Este compuesto mostró una actividad
anticoccidial significativa para todos los parámetros estudiados (mortalidad,

16
aumento de peso, conversión alimenticia, puntaje de caída y puntaje de lesión).
La mortalidad debida a la coccidiosis se redujo al 0,1 % en las aves infectadas
con medicamentos.
Baghlaf et al., (1979) obtuvieron mediante una fermentación que contenía los
siguientes ingredientes: melaza negra, levadura forrajera, salvado de arroz,
KH2PO4 y agua del grifo, una producción de oxitetraciclina por Streptomyces
rimosus 1207, como suplemento alimenticio para animales. Al puré seco que
contenía la oxitetraciclina se le analizó los aminoácidos l-arginina, l-histidina, l-
lisina, l-isoleucina, l-leucina, l-fenilalanina, dl-metionina y l-triptófano, los cuales
podrían ser de valor nutricional para pollos.
En otro estudio Pensack et al., (1982), determinaron los efectos de la
administración de avoparcina, un antibiótico glucopéptido producido por la
fermentación de una cepa de Streptomyces candidus, a pollos de engorde en
Taiwán, Australia, Europa y los Estados Unidos. La avoparcina mejoró
significativamente el peso vivo y la eficacia de la alimentación a los niveles de la
dieta probada (5,5 a 22 ppm). La avoparcina en la alimentación fue eficaz para
aumentar la tasa de ganancia de peso y mejorar la eficiencia de la alimentación
de los pollos de engorde.
Por otro lado George et al., (1982a) analizaron veinte pollos de engorde
alimentados con bambermicinas (Flavomicina, un antibiótico producido por
Streptomyces). Las aves fueron inoculadas (dosificadas) el día diez y 11 de
alimentación con Salmonella typhimurium. El estudio evaluó los efectos de las
bambermicinas sobre la incidencia de Salmonella, el desprendimiento y la
resistencia a los antimicrobianos. Las bambermicinas no tuvieron efecto sobre el
peso corporal. La duración del desprendimiento de salmonelas, el número de
salmonelas arrojados el día tres después de la dosificación, la capacidad de
recuperación del tejido de las salmonelas y el número total de patrones de
resistencia a bambermicina dio lugar a la disminución de salmonelas.
George et al., (1982b) realizaron ensayos con pollos de engorde machos para
evaluar los efectos de la virginiamicina como tratamiento contra la infección por
enteritis necrótica inducida experimentalmente. Las aves se dosificaron por vía
oral con 10 mL de cultivo de Clostridium perfringens. Las aves alimentadas con
virginiamicina tuvieron significativamente menos mortalidad y menores
puntuaciones de lesiones intestinales que las aves no medicadas cuando se

17
infectaron experimentalmente con enteritis necrótica. Aunque los promotores del
crecimiento antibiótico en la dieta se han usado durante mucho tiempo para
aumentar el rendimiento del crecimiento en la producción comercial de alimentos
para animales, los detalles bioquímicos asociados con estos efectos permanecen
pobremente definidos.
Gadde et al., (2018) usaron un enfoque metabólico para caracterizar e identificar
los compuestos bioquímicos presentes en el intestino de pollos de engorde
alimentados con una dieta estándar no suplementada y una dieta suplementada
con promotores del crecimiento (antibióticos: virginiamicina o bacitracina y
disalicilato de metileno). En los pollos suplementados con virginiamicina y con
bacitracina, los cambios en los niveles bioquímicos e intestinales proporcionaron
un patrón bioquímico distintivo única para cada grupo suplementado con estos
antibióticos. Estas características bioquímicas se caracterizaron por aumentos en
los niveles de metabolitos de aminoácidos (por ejemplo, 5-hidroxilisina, 2-
aminoadipato, 5-hidroxiindoleaceato, 7-hidroxiindol sulfato), ácidos grasos (por
ejemplo, oleato, vaccenato, eicosapentaenoato, 16-hidroxi-palmitato, estearato),
nucleósidos (por ejemplo, inosina, N6-metiladenosina) y vitaminas (por ejemplo,
nicotinamida). Estos resultados proporcionan el marco para estudios futuros para
identificar compuestos químicos naturales para mejorar el rendimiento de
crecimiento avícola sin el uso de antibióticos en la alimentación.
Xu et al., (2018) en una investigación donde utilizan Bacillus licheniformis como
agente normalizador de la microbiota del íleon de pollos infectados con enteritis
necrótica, encontraron que la abundancia de Streptomyces spp., fue mayor que
que otros grupos. Lo cual plantea que los Streptomyces spp., tienen redes
reguladoras de metabolismo secundarias complejas y grandes, pueden producir
muchas sustancias bioactivas beneficiosas a través del metabolismo secundario.
La estreptomicina es uno de los metabolitos secundarios producido por
Streptomyces, el cual tiene una potente actividad antibacteriana contra
Mycobacterium tuberculosis y eficacia antimicrobiana contra varias bacterias
Gram negativas (por ejemplo, E. coli, Salmonella y Brucella). Además de los
antibióticos, el metabolismo secundario de Streptomyces spp., es variado, ya que
el mismo incluye agentes antitumorales, inhibidores inmunitarios, agentes de
resistencia a insectos y enzimas exocelulares (por ejemplo, pectinasa, celulasa y
quitinasa). Además, investigaciones recientes sugieren que Streptomyces spp.,

18
podría tener beneficios que se extienden más allá del intestino, como curar la
Salmonella pullorum y Pasteurella multocida en pollos infectados (Xu et al., 2018).

19
CAPÍTULO 2. ESTUDIO DE LA
TOLERANCIA DE LA CEPA
STREPTOMYCES sp. RL8 A
SALES BILIARES, CLORURO DE
SODIO Y pH

20
CAPÍTULO 2. ESTUDIO DE LA TOLERANCIA DE LA CEPA STREPTOMYCES
sp. RL8 A SALES BILIARES, CLORURO DE SODIO Y pH
Introducción
La Organización de las Naciones Unidas para la Agricultura y Alimentos
(FAO/WHO) y la Organización Mundial de la Salud (OMS), establecieron en 2002
criterios de selección de microorganismos probióticos en el “Informe del grupo de
trabajo sobre la redacción de directrices para la evaluación de los probióticos en
los alimentos” (WHO, 2002). Uno de los criterios de selección in vitro es la
resistencia a la acidez estomacal y a las sales biliares de intestino (Park et al.,
2006, Cueto-Vigil et al., 2010).
La especificidad de especies probióticas en el animal, es un factor importante que
interfiere en la colonización y en la adhesión in vivo de los microorganismos
(Frizzo et al., 2006); es necesario realizar una adecuada evaluación de cepas de
acuerdo con diferentes criterios de selección, (resistencia a la acidez, sales,
temperatura, entre otras), de forma tal que los microorganismos colonizadores
lleguen en estado viable y en cantidades suficientes una vez que han superado
las barreras ácida y biliar en el tracto digestivo (Tuomola et al., 2001); los
resultados de estas pruebas también pueden predecir la capacidad in vivo de las
cepas; de ahí que, el éxito de un probiótico depende en gran medida de realizar
una buena selección in vitro (Chichlowski et al., 2007).
Por lo antes expuesto el siguiente capítulo tiene como objetivos:
1. Determinar la tolerancia de la cepa RL8 a diferentes valores de pH.
2. Determinar la tolerancia de la cepa RL8 a diferentes concentraciones de
sales biliares.
3. Determinar la tolerancia de la cepa RL8 a diferentes concentraciones de
cloruro de sodio.
2.1 Materiales y Métodos
2.1.1 Determinación de la tolerancia a diferentes valores de pH
Se evaluó la cepa RL8 a diferentes valores de pH 2, 3, 4, 5, 6 y 7 (incubación a 25
°C durante una semana); la sobrevivencia y resistencia se comprobó al comparar
el conteo de microorganismos viables del inóculo, con las células sobrevivientes
después de la incubación a diferentes valores de pH en caldo triptona soya según
Bernal, (2016); el porcentaje de resistencia fue calculado por la siguiente ecuación
(Rondón et al., 2009):

21
% R pH = [(Cel/mL) TSB pH x 100] / (Cel/mL) TSB pH 7 (inóculo)
El inóculo inicial para cada tolerancia fue de 1.5 x108 Cel/mL.
2.1.2 Determinación de la tolerancia a diferentes concentraciones de sales
biliares
El ensayo fue realizado a diferentes concentraciones de sales biliares 0,05; 0,1,
0,15; 0,3; 0,4 y 0,5 % (p/v) ajustado a pH 7 con HCI 5N y NaOH 5N en medio de
cultivo caldo triptona soya por sus siglas en inglés (TSB), con un inóculo a una
concentración de 1.5 x108 Cel/mL (incubación a 25 °C durante siete días) (Bernal
et al., 2016); al cabo de este tiempo la sobrevivencia y resistencia a sales biliares
se determinó mediante la ecuación según Rondón et al., (2009):
% R= [(Cel/mL) TSB + Sales Biliares x 100] / (Cel/mL) TSB (inóculo)
2.1.3 Determinación de la tolerancia a diferentes concentraciones de cloruro
de sodio
La prueba se realizó utilizando TSB y diferentes concentraciones de NaCl, 2, 4, 7,
9 y 10 % (p/v), (incubación a 25 °C durante siete días) (Bernal, 2016); finalizado el
tiempo de incubación se determinó el crecimiento de la RL8 a altas concentra-
ciones de NaCl, se utilizó la ecuación de Rondón et al., (2009).
% R= [(Cel/mL) TSB + NaCl x 100] / (Cel/mL) TSB (inóculo)
2.2 Resultados y Discusión
2.2.1 Determinación de la tolerancia de la cepa RL8 a diferentes valores de
pH
La cepa evaluada a pH 2 no mostró buenos resultados durante el transcurso de la
semana que duró el ensayo (Fig. 1). Para contrarrestar este resultado negativo,
se realizó un nuevo experimento (Fig. 2) donde se avaluó la misma cepa RL8 a
pH 1, 2 y 3 en el transcurso de seis horas tomándose muestras cada dos hora, se
sembró la dilución 103 y 104 en agar caseína almidón (ACA) similar a Rangel,
(2015) para evaluar en el tiempo la viabilidad de la cepa de Streptomyce sp. RL8
en UFC/mL según Rondón, (2008), ya que según Vergara et al., (1989)
observaron que los tiempos de retención de los alimentos en pollos hasta los 21
días de edad es aproximadamente de 4 horas y 51 minutos, y según Angel et al.,
(2013), los pH en el proventrículo y molleja suelen estar a edades tempranas de 1
día a 21 día desde 1,9-2,9 a 2,8-4,6 respectivamente.
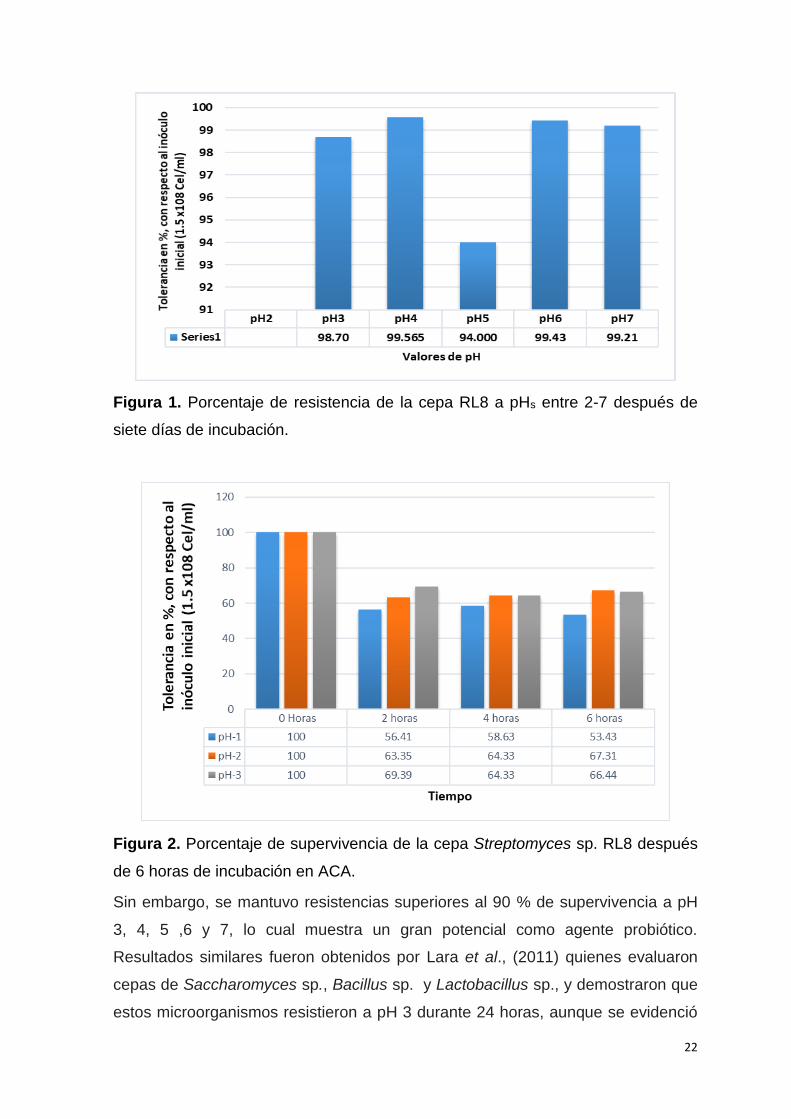
22
Figura 1. Porcentaje de resistencia de la cepa RL8 a pHs entre 2-7 después de
siete días de incubación.
Figura 2. Porcentaje de supervivencia de la cepa Streptomyces sp. RL8 después
de 6 horas de incubación en ACA.
Sin embargo, se mantuvo resistencias superiores al 90 % de supervivencia a pH
3, 4, 5 ,6 y 7, lo cual muestra un gran potencial como agente probiótico.
Resultados similares fueron obtenidos por Lara et al., (2011) quienes evaluaron
cepas de Saccharomyces sp., Bacillus sp. y Lactobacillus sp., y demostraron que
estos microorganismos resistieron a pH 3 durante 24 horas, aunque se evidenció

23
una disminución porcentual del crecimiento en los tres géneros. Sin embargo, a
pH 4 se presentó el mayor porcentaje de crecimiento 70, 90 y 95 %, lo cual indicó
una supervivencia en condiciones de acidez; mientras que a pH 5, 6 y 7
disminuyó gradualmente, pero también se mantuvo estable el crecimiento, no
siendo así con la cepa de Streptomyces sp. RL8 evaluada en esta investigación la
cual obtuvo valores de 98,7 % y 99,56 % de supervivencia a pH 3 y 4, y 94 % y
99,21 % a pH 5 y pH 7. Bernal et al., (2015) y Prakashwadekar et al., (2015) en
una evaluación con este mismo género de microorganismo no obtuvieron
crecimiento a valores de pH de 1-3, pero crecieron a un pH superior a tres. Se ha
estimado que la tasa de supervivencia de los probióticos tradicionales en el
intestino del huésped es solo del 20-40 %, siendo la acidez gástrica uno de los
principales obstáculos para que los probióticos ejerzan su acción (Bezkorovainy,
2001). A pesar de que esta cepa de actinomiceto no crece a pH entre 1 y 2, en un
tiempo prolongado, se pueden esperar mayores tasas de supervivencia en
comparación con los probióticos bacterianos tradicionales, ya que los
actinomicetos son capaces de producir esporas resistentes a condiciones
adversas como la acidez (Bernal et al., 2015).
Introducir como criterio de selección la supervivencia a pHs menores de 3,
permite detectar a los microorganismos resistentes a estas condiciones extremas,
sobre todo si se parte del hecho de que el jugo gástrico de los pollos puede llegar
a tener valores de pH entre 0,5 - 2,0 (Ehrmann et al., 2002)
2.2.2 Determinación de la tolerancia de la cepa RL8 a diferentes
concentraciones de sales biliares
En esta investigación los valores de supervivencia en TSB enriquecido con sales
biliares a pH 7 fue por encima del 100 % (Fig.3). Los valores más bajos de
resistencia fueron encontrados a concentraciones de sales biliares de 0,05 % (p/v)
(62,5 %), 0,1 % (p/v) (1,11) y 0,15 % (p/v) (4,29). Los valores de resistencia más
altos fueron a 0,3 % (p/v) (157,89 %), 0,4 % (p/v) (142 %) y 0,5 % (p/v) (125 %).
De acuerdo a los resultados obtenidos se puede decir que la cepa de
Streptomyces sp. RL8 es capaz de sobrevivir a concentraciones de sales biliares
desde 0,05 % (p/v) hasta 0,5 % (p/v), pudiendo desarrollar sus actividades
metabólicas sin verse completamente inhibida. Esta distinción es primordial si las
cepas deben usarse como probióticos porque deberían estar creciendo
activamente para ejercer su efecto. Estos resultados son similares a los obtenidos

24
por Bernal et al., (2015) cuando analizaron la cepa RL8 y otras cuatro cepas
pertenecientes al mismo género (La7, La12, N7 y V4), al evaluar su posible
acción como agente probiótico en la acuicultura. Según Begley et al., (2005) los
microorganismos comensales y patógenos deben resistir las acciones
perjudiciales de la bilis para sobrevivir en el intestino de los animales.
Figura 3. Tolerancia de la cepa RL8 a diferentes concentraciones de sales
biliares después de siete días de incubación.
2.2.3 Determinación de la tolerancia de la cepa RL8 a diferentes
concentraciones de cloruro de sodio
En la (Fig.4) se muestra la tolerancia (%) de la cepa RL8 a diferentes
concentraciones de NaCl. Como se puede observar hubo una alta tendencia al
crecimiento de la cepa RL8 bajo estas condiciones, lo cual es altamente
significativo comparado con otros grupos de microorganismos candidatos a
probióticos. Esto es posible debido a que la cepa de Streptomyces sp. RL8 es
aislada de sedimentos marinos por Bernal et al., (2015) lo cual corrobora los
resultados obtenidos.
La mayoría de las bacterias son relativamente insensibles a las variaciones de la
presión osmótica, y se adaptan a cambios fuertes con respecto a la concentración
de solutos del medio, porque poseen una pared celular mecánicamente rígida
(Voigt, 2003). Basados en lo anterior, se deduce que la cepa de Streptomyce sp.
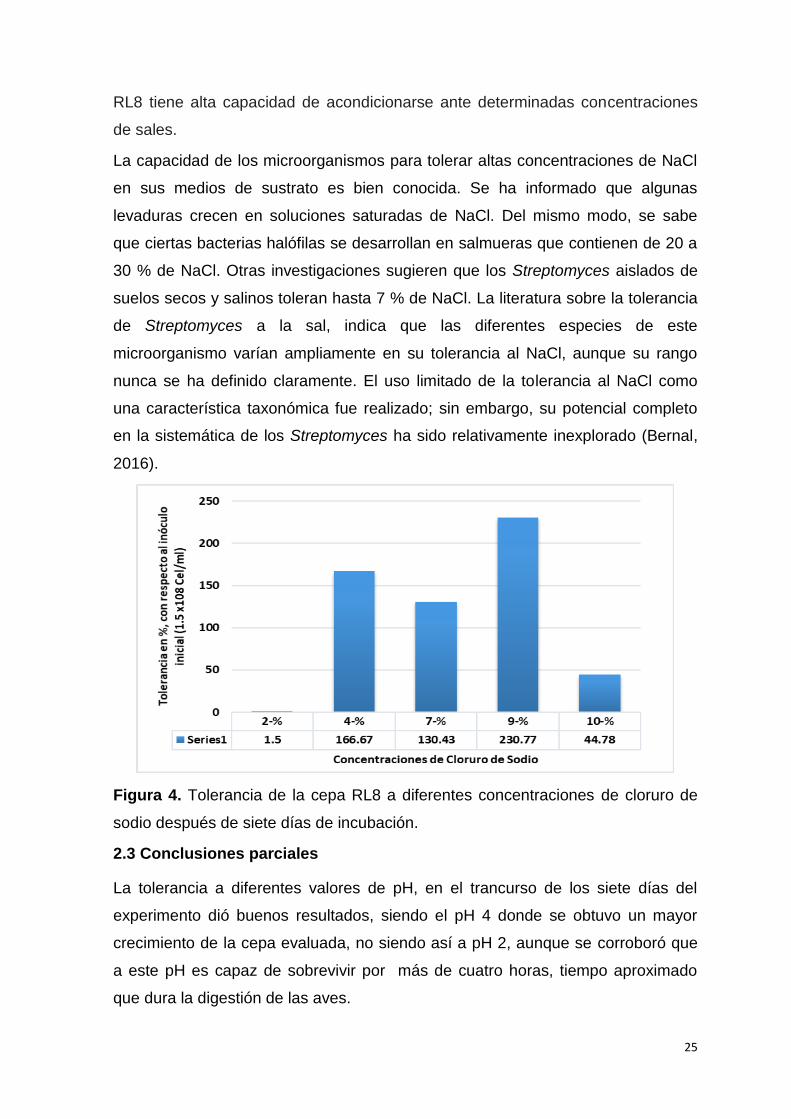
25
RL8 tiene alta capacidad de acondicionarse ante determinadas concentraciones
de sales.
La capacidad de los microorganismos para tolerar altas concentraciones de NaCl
en sus medios de sustrato es bien conocida. Se ha informado que algunas
levaduras crecen en soluciones saturadas de NaCl. Del mismo modo, se sabe
que ciertas bacterias halófilas se desarrollan en salmueras que contienen de 20 a
30 % de NaCl. Otras investigaciones sugieren que los Streptomyces aislados de
suelos secos y salinos toleran hasta 7 % de NaCl. La literatura sobre la tolerancia
de Streptomyces a la sal, indica que las diferentes especies de este
microorganismo varían ampliamente en su tolerancia al NaCl, aunque su rango
nunca se ha definido claramente. El uso limitado de la tolerancia al NaCl como
una característica taxonómica fue realizado; sin embargo, su potencial completo
en la sistemática de los Streptomyces ha sido relativamente inexplorado (Bernal,
2016).
Figura 4. Tolerancia de la cepa RL8 a diferentes concentraciones de cloruro de
sodio después de siete días de incubación.
2.3 Conclusiones parciales
La tolerancia a diferentes valores de pH, en el trancurso de los siete días del
experimento dió buenos resultados, siendo el pH 4 donde se obtuvo un mayor
crecimiento de la cepa evaluada, no siendo así a pH 2, aunque se corroboró que
a este pH es capaz de sobrevivir por más de cuatro horas, tiempo aproximado
que dura la digestión de las aves.

26
La resistencia a sales biliares mantuvo un buen crecimiento, aunque las
concentraciones que más se destacaron fueron 0.3 % (p/v) (157.89 % de
resistencia), 0.4 % (p/v) (142 % de resistencia) y 0.5 % (p/v) (125 % de
resistencia) respectivamente.
Como era de esperar este microorganismo marino es capaz de soportar altas
concentraciones de NaCl, lo cual fue corroborado con la siguiente investigación
donde se obtuvieron tolerancias superiores a 100 %, lo cual indica su excelente
resistencia a altas concentraciones de cloruro de sodio.

27
CAPÍTULO 3.
EVALUACIÓN DEL
EFECTO DE LA CEPA
STREPTOMYCES sp. RL8
SOBRE LOS
PARÁMETROS
BIOPRODUCTIVOS Y DE
SALUD

28
CAPÍTULO 3. EVALUACIÓN DEL EFECTO DE LA CEPA STREPTOMYCES sp.
RL8 SOBRE LOS PARÁMETROS BIOPRODUCTIVOS Y DE SALUD
Introducción
Generalmente, en la evaluación in vivo se realizan estudios de supervivencia y
persistencia en el TGI, desafío experimental con microorganismos patógenos y
determinación de propiedades inmunomoduladoras en el hospedero a través de
diversas metodologías. En este sentido, Milián, (2009) y Rondón, (2009)
desarrollaron métodos para la evaluación multifactorial de aditivos probióticos en
pollos de ceba que incluyen la determinación de indicadores microbiológicos,
morfométricos, hematológicos, inmunológicos, bioproductivos y de salud (García,
2011).
En el capítulo 2 de la presente tesis se demostró las características in vitro de la
cepa de Streptomyces sp. RL8 como candidata a agente probiótico. Sin embargo,
es imprescindible comprobar la respuesta biológica de los animales cuando se
adicionan estos microorganismos. Por tal motivo el objetivo de este capítulo es:
1. Evaluar la actividad probiótica de la cepa de Streptomyces sp. RL8 sobre
los indicadores fisiológicos y bioproductivos de las aves evaluadas hasta los 21
días de edad.
3.1 Materiales y Métodos
Animales y dieta basal
En el experimento se utilizaron 60 animales 30 hembras y 30 machos de la raza
Leghorn (1 día de edad, con peso vivo promedio de 38,72 g), los mismos fueron
distribuidos a razón de 30 aves por tratamiento (Fig.5). Los animales recibieron el
agua ad libitum y el alimento a base de maíz y soya enriquecido (Anexo 1) con el
suplemento probiótico 108 UFC·g-1 (García, 2011).
Microorganismos: Streptomyces sp. RL8
La cepa RL8 se preparó semanalmente en TSB, se mezcló con el pienso y se
conservó a 25 ºC hasta su utilización. Sustrato sólido CAR de subproducto de la
producción agrícola.
Tratamientos experimentales
Se establecieron dos grupos de animales en función de los objetivos. Las aves se
trataron hasta los 21 días de edad. Cada cultivo se mezcló diariamente de forma
manual con la dieta basal. Los grupos experimentales fueron:

29
Grupo control (I): sin aditivo.
Grupo RL8: con adición de la cepa de Streptomyces sp. RL8 a una concentración
de 108 UFC·g-1.
Condiciones experimentales y sistema de manejo de los animales
Los muestreos se realizaron a los 21 días de edad de los animales, se alojaron en
jaulas metálicas y se utilizó una densidad de 25 aves m2 hasta los 18 días de
edad, posteriormente se redistribuyeron según el tratamiento a 12-13 aves m2.
El sistema de vacunación de los animales consistió en una dosis por aspersión de
Marek y Gumboro el primer día de edad.
Procedimiento experimental para la toma y análisis de las muestras
Se seleccionaron al azar quince animales de cada tratamiento, en cada muestreo,
se pesaron y se sacrificaron por desangramiento de la vena yugular descrito por
Sánchez, (1990) citado por García, (2011).
Se tomó muestras de 1 mL de sangre en tubos con 5 µl heparina sódica. La
actividad probiótica de los microorganismos se determinó en función de los
siguientes indicadores: Peso vivo, Ganancia media diaria (GMD), Incremento de
peso (Byrd et al. 2003), peso relativo de pechuga y piernas. Para analizar los
parámetros morfométricos se pesaron, intestino delgado (ID), intestino grueso
(IG), los ciegos vacíos, hígado y pechuga. Se determinó el peso del bazo como
órgano inmune, en una balanza técnica Sartorius BL 1 500. El peso relativo de
estos órganos se expresó en g kg-1 de peso vivo del animal (García, 2011).
Análisis estadístico
Los datos se procesaron mediante análisis de varianza según diseño
completamente aleatorizado. Antes de aplicar el ANOVA se procedió a verificar la
normalidad de los datos mediante la prueba de Shapiro Wilk y para la
homogeneidad de la varianza se utilizó la prueba de Levene (Sokal y Rohlf, 1995).
Para detectar diferencias significativas en los valores de crecimiento en función
del tratamiento con la cepa de Streptomyces sp. RL8 suministrada y el grupo
control, se empleó la prueba de comparación de Duncan, (1955). Para todos los
análisis efectuados, el nivel de significación fue de p<0,05. Los análisis
estadísticos se realizaron utilizando el software STATISTICA versión 8.0 para
Windows.

30
Figura 5. Diagrama para la evaluación de la actividad probiótica de Streptomyces
sp. RL8 en pollos de remplazo Leghorn a los 21 días de edad (Adaptado de
Rondón, 2009 y García, 2011).
3.2 Resultados y Discusión
Productivos y de Salud
La tabla uno, muestra el comportamiento de los indicadores bioproductivos de los
animales durante la evaluación del microorganismo candidato a probiótico.
Estadísticamente no hubo diferencias significativas entre el grupo tratado con la
cepa de Streptomyces sp. RL8 y el control. Los valores obtenidos en este
experimento fueron muy inferiores a los que obtuvo Gómez, (2013) y García,
(2011), al tratar pollos de ceba con Lactobacillus spp. Todo parece indicar que el
probiótico utilizado no ejerció efecto sobre el peso de los pollos evaluados.

31
Tabla 1: Pesos relativos de los órganos (g*Kg-1) a los 21 días de tratamiento.
Indicadores
Tratamientos Peso Inicial
(g)
1d
Peso Final (g)
21d
Incremento
de Peso
(g) a los 21d
Ganancias
Media Diaria
(g) a los 21 d
RL8-H 39,07 108,38 69,31 3,30
RL8-M 38,56 86,93 48,37 2,30
RL8-T 38,82 97,65 58,83 2,80
Control-H 38,86 108,33 69,47 3,30
Control-M 38,32 98,146 59,82 2,84
Control-T 38,59 103,24 64,65 3,07
EE± 0,50 18,02 7,98 0,38
a,b,c, Por columna las medias con diferentes letras difieren a p<0,05 (Duncan, 1955). RL8-H: Media de
los Tratamientos con la Cepa RL8 en pollos en Hembras, RL8-M: Media de los Tratamiento con la
Cepa RL8 en pollos Machos, RL8-T: Media Total del tratamiento con la cepa RL8, Control-H: Grupo
control Hembra sin probiótico, Control-M: Grupo Control Macho sin probiótico, Control-T: Media total
del grupo control sin probiótico, EE±: Error estándar de la media.
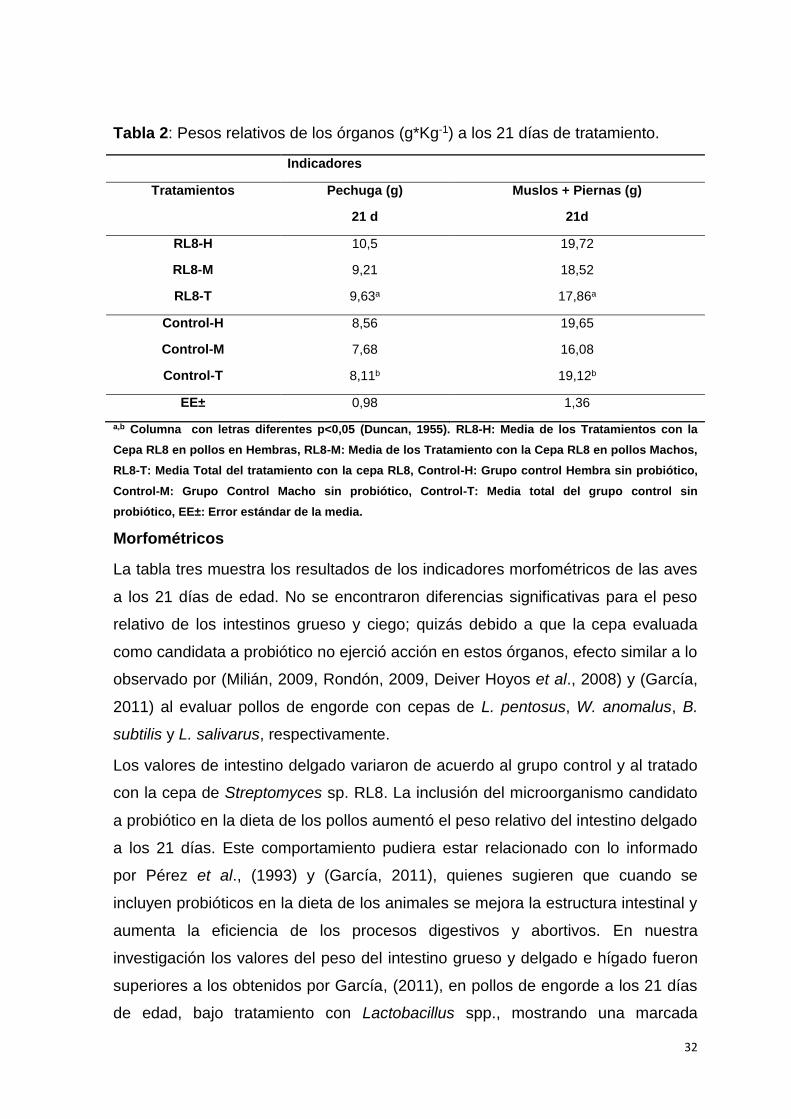
Los parámetros bioproductivos y de salud mostrados en la tabla 2, peso de
pechuga (g) y muslos más piernas (g), no mostraron diferencias estadísticamente
significativas entre el grupo tratado con Streptomyces sp. RL8 y el grupo control.
Los mayores pesos de la pechuga se encontraron en el grupo tratado con la cepa
de Streptomyces sp. RL8, mientras que los mayores valores encontrados en
cuanto al peso de los muslos más piernas fueron mostrados por el grupo control.
Es posible que los microorganismos candidatos a probióticos evaluados actúen en
el metabolismo del animal, tanto energético como proteico, al obtenerse un estado
eubiósico en el intestino, acción que da lugar al aumento del tiempo de recambio
de las células intestinales, lo que provoca ahorro sustancial en los nutrientes
digeridos por el animal y posibilita que estos puedan utilizarse en la producción de
masa muscular, entre otras funciones (García, 2011). Es importante destacar que
se evaluaron gallinas ponedoras de la raza Leghorn, las cuales no se caracterizan
por una elevada producción de carne, su importancia económica radica en la
puesta de huevos, lo cual no fue objeto de estudio en esta investigación.
32
Tabla 2: Pesos relativos de los órganos (g*Kg-1) a los 21 días de tratamiento.
Indicadores
Tratamientos Pechuga (g)
21 d
Muslos + Piernas (g)
21d
RL8-H 10,5 19,72
RL8-M 9,21 18,52
RL8-T 9,63a 17,86a
Control-H 8,56 19,65
Control-M 7,68 16,08
Control-T 8,11b 19,12b
EE± 0,98 1,36
a,b Columna con letras diferentes p<0,05 (Duncan, 1955). RL8-H: Media de los Tratamientos con la
Cepa RL8 en pollos en Hembras, RL8-M: Media de los Tratamiento con la Cepa RL8 en pollos Machos,
RL8-T: Media Total del tratamiento con la cepa RL8, Control-H: Grupo control Hembra sin probiótico,
Control-M: Grupo Control Macho sin probiótico, Control-T: Media total del grupo control sin
probiótico, EE±: Error estándar de la media.
Morfométricos
La tabla tres muestra los resultados de los indicadores morfométricos de las aves
a los 21 días de edad. No se encontraron diferencias significativas para el peso
relativo de los intestinos grueso y ciego; quizás debido a que la cepa evaluada
como candidata a probiótico no ejerció acción en estos órganos, efecto similar a lo
observado por (Milián, 2009, Rondón, 2009, Deiver Hoyos et al., 2008) y (García,
2011) al evaluar pollos de engorde con cepas de L. pentosus, W. anomalus, B.
subtilis y L. salivarus, respectivamente.
Los valores de intestino delgado variaron de acuerdo al grupo control y al tratado
con la cepa de Streptomyces sp. RL8. La inclusión del microorganismo candidato
a probiótico en la dieta de los pollos aumentó el peso relativo del intestino delgado
a los 21 días. Este comportamiento pudiera estar relacionado con lo informado
por Pérez et al., (1993) y (García, 2011), quienes sugieren que cuando se
incluyen probióticos en la dieta de los animales se mejora la estructura intestinal y
aumenta la eficiencia de los procesos digestivos y abortivos. En nuestra
investigación los valores del peso del intestino grueso y delgado e hígado fueron
superiores a los obtenidos por García, (2011), en pollos de engorde a los 21 días
de edad, bajo tratamiento con Lactobacillus spp., mostrando una marcada

33
diferencia entre ambos grupos tratados con Streptomyces sp. RL8. Efecto similar
se evidenció en los ciegos de los animales tratados donde mostraron valores
superiores a los obtenidos por (García, 2011).
En el estudio realizado por García, (2011), los ciegos no variaron su peso, estos
variaron después de los 35 días de edad, donde se incluyó L. pentosus LB-31 y
su mezcla con W. anomalus LV-6 al compararlos con un grupo control. Los
factores que pueden influir en el tamaño de estos órganos están relacionados con
la dieta y los procesos metabólicos celulares y microbianos que aquí ocurren
(Chichlowski et al., 2007).
Tabla 3: Pesos relativos de los órganos (g*Kg-1) a los 21 días de tratamiento.
Tratamientos Intestino
Delgado
21 días
Intestino
Grueso
21 días
Ciegos
21 días
Hígado
21 días
Bazo
RL8-H 98,94 0,98 11,13 38,19 2.35
RL8-M 101,94 0,76 11,31 40,31 2.23
RL8-T 100,46a 0,87a 11,22a 39,25a 2.29a
Control-H 88,45 0,85 8,88 37,04 1.81
Control-M 92,96 1,036 12,31 37,157 1.72
Control-T 90,71b 0,94a 10,59a 37,10b 1.77b
EE± 10,82 0,12 1,34 2,73 0.29
a,b,c, Por columna las medias con diferentes letras difieren a p<0,05 (Duncan, 1955). RL8-H: Media de
los Tratamientos con la Cepa RL8 en pollos en Hembras, RL8-M: Media de los Tratamiento con la
Cepa RL8 en pollos Machos, RL8-T: Media Total del tratamiento con la cepa RL8, Control-H: Grupo
control Hembra sin probiótico, Control-M: Grupo Control Macho sin probiótico, Control-T: Media total
del grupo control sin probiótico, EE±: Error estándar de la media.
3.3 Conclusiones parciales
La inclusión de la cepa de Streptomyces sp. RL8 en el alimento de los pollos no
modificó los indicadores bioproductivos de los animales con respecto al grupo
control, lo cual indica que el probiótico utilizado no ejerce efecto sobre el peso de
los pollos evaluados.
La utilización de la cepa de Streptomyces sp. RL8 incrementó el peso relativo del
hígado, intestino delgado, intestino grueso, bazo y ciego; quizás debido a que los
aditivos microbianos evaluados ejercieron acción en estos órganos.

34
CAPÍTULO 4. ESTUDIO DE LOS
INDICADORE HEMATOLÓGICOS AL
EVALUAR LA CEPA DE
STREPTOMYCES sp. RL8 Y POSIBLE
EFECTO MODULADOR DEL SISTEMA
INMUNE EN POLLOS RAZA LEGHORN

35
CAPÍTULO 4. ESTUDIO DE LOS INDICADORES HEMATOLÓGICOS AL
EVALUAR LA CEPA DE STREPTOMYCES sp. RL8 Y SU POSIBLE EFECTO
MODULADOR DEL SISTEMA INMUNE EN POLLOS RAZA LEGHORN
Introducción
Los signos clínicos en las aves son generalmente inespecíficos y la información
obtenida por el examen físico es limitada. La evaluación de muestras de sangre
puede servir como herramienta importante para el seguimiento de la salud de las
aves, en el diagnóstico de enfermedades, durante los tratamientos y para
observar las condiciones de salud del animal. La sangre es esencial para el
mantenimiento de electrolisis y del agua, para el control de la temperatura y para
el buen funcionamiento del sistema inmune. La utilización más usual de la
hematología es para monitorear la salud general del animal, evaluar su capacidad
para transportar oxígeno y defenderse contra los agentes infecciosos (Voigt,
2003). La bolsa de Fabricio y el bazo junto al timo, las tonsilas cecales, las
glándulas de Harder y la médula ósea son los órganos responsables de la
inmunidad en las aves, que durante los primeros días del crecimiento del animal
reciben más nutrientes en comparación con los tejidos musculares (Contreras &
Fernández, 1999, Mateos et al., 2002., Perozo-Marín et al., 2004). Así cuando los
animales llegan a la madurez sexual, la bolsa de Fabricio y el timo involucionan y
la respuesta inmune pasa a depender del sistema periférico (García, 2011).
El objetivo de este capítulo estuvo encaminado a:
1. Evaluar la actividad probiótica de la cepa de Streptomyces sp. RL8 en los
indicadores hematológicos, y su posible efecto modulador sobre el sistema
inmune de las aves evaluadas.
4.1 Materiales y Métodos
A las muestras de sangre (1 mL de cada animal en 5 µL de EDTA) se le
determinó la hemoglobina (Hb) (g/L), hematocrito (Ht) (L/L) y Concentración de
Hemoglobina Corpuscular Media (CHCM) (g/L), linfocitos (1/L), glóbulos blancos
(x109), heterófilos (1/L) y eosinófilos (1/L) según Sadeghi et al., (2015). Para la
determinación de hemoglobina se utilizó el método de la cianometahemoglobina
descrito por Crosby et al., (1954), para ello se tomó 0,2 mL de sangre diluida y se

36
mezcló con 5 mL del reactivo de Drabkin y después de 10 minutos se realizó la
lectura de la absorbancia en un equipo marca GENESIS 6 de la Thermo Electron
Corporation® a 540 nm y se usó el agua destilada como blanco.
Para la medición del hematocrito se realizó el llenado de sangre de los capilares
para microhematocrito hasta una tercera parte del capilar. Estos se sellaron con
mechero y se centrifugaron a 12000 rpm (equivalente a 16128 fuerza centrífuga
relativa (FCR)) durante diez minutos en una microcentrífuga marca MLW modelo
TH21. Posteriormente, se realizó la lectura en equipo de microhematocrito marca
Hawkley con lector de escala móvil que permitió situarlo en el nivel de los
sedimentos hemáticos y proceder entonces a la lectura similar a (García, 2011).
Tratamiento estadístico
Los datos se procesaron según tratamiento estadístico utilizado en el capítulo III.
Para detectar diferencias significativas en los valores de crecimiento en función
del tratamiento con la cepa de Streptomyces sp. RL8 suministrada y el grupo
control, se empleó la prueba de comparación de Duncan, (1955). Para todos los
análisis efectuados, el nivel de significación fue de p<0,05. Los análisis
estadísticos se realizaron utilizando el software STATISTICA versión 8.0 para
Windows.
4.2 Resultados y Discusión
La tabla tres muestra los resultados del análisis hematológico. Aunque los valores
de hematocrito (Hto) y hemoglobina (Hb) se mantuvieron dentro de los rangos
normales de crianza (0,23-0,55 L/L y 70-186 g/L, respectivamente (Perozo-Marín
et al., 2004)) en ambos grupos, este último parámetro fue significativamente
superior en el grupo tratado con Streptomyces sp. RL8, con respecto al grupo
control. Sin embargo, estos valores de hemoglobina y hematocrito fueron
inferiores a los que obtuvo (Mchugh, 2002) en pollos de ceba, pero coinciden con
los obtenidos por (García, 2011) al evaluar L. pentosus LB-31, W. anomalus LV-6
y su mezcla en pollos de engorde; y con Mascheck & Baker., (2008) quienes
evaluaron pollos de ceba tratados con Saccharomyces cerevisiae, L. acidophilus,
B. subtilis y una mezcla de los tres.
Haile & Chanie, (2014) plantearon que los pollos suelen tener valores de
hematocrito relativamente bajos (tan bajo como 0,24 L/L), los cuales aumentan
con la edad. Los resultados difieren de los encontrados por Valderr ama, (2013)

37
con valores medios de hematocrito del 0,36 L/L en pollos de engorde alimentados
con probióticos a base de Lactobacillus, Streptococcus y Bifidobacterium.
Igualmente, Mchugh, (2002) encontraron valores de hematocrito de 0,36 L/L en
pollos de engorde suplementados con B. subtilis. Según Mascheck & Baker.,
(2008), es probable que el tipo de dieta, las proporciones de los nutrientes
utilizados, y la raza, pudieran influir en los resultados discrepantes.
El conteo de glóbulos blancos no difirió entre el grupo tratado y el control, y se
mantuvo dentro de los valores normales para aves. Sin embargo, fueron inferiores
a los que obtuvieron Rahimi, (2006) al tratar pollos de engorde con el antibiótico
virgianiamycim y el probiótico comercial BioPlus 2B®, y similares a los alcanzados
por Sadeghi et al., (2015) al tratar pollos de ceba con Gallipro®.
No se detectaron diferencias significativas entre el grupo control y el tratado con la
cepa de Streptomyces sp. RL8 en cuanto al conteo linfocitario; cuyo valor estuvo
dentro del rango del 0,68-0,77 (1/L) obtenido por Sadeghi et al., 2015, Areces &
Céspedes, 1992) quienes usaron el probiótico comercial Gallipro®. Por el
contrario, Areces & Céspedes, (1992) obtuvieron valores inferiores a los
alcanzados en este trabajo; al realizar un estudio con pollos ponedores de la raza
Hy-Line W-98, utilizando dietas enriquecidas con enzimas, probióticos y
combinaciones entre ambos.
Tabla 3: Resultados del análisis hematológico.
Tratamientos Hb (g/L) Hto (L/L) GB x 109/L L (1/L) H (1/L) E (1/L) CHCM (g/L)
RL8 88a 0,26a 1,79a 0,76a 0,21a 0,09a 333,0
Control 82a 0,24b 1,72a 0,76a 0,20b 0,03b 333,0
EE+ 1,7 0,51 0,87 2,7 2,6 2,22 0
a, b, Dentro de la misma columna, Indica diferencias entre las medias para un valor de p<0,05
(Bradford, 2000). Hb-Hemoglobina, Hto-Hematocrito, GB-Glóbulos Blancos, L-Linfocitos, H-
Heterófilos, E-Eosinófilos, CHCM-Hemoglobina corpuscular media.
Los heterófilos son leucocitos polimorfonucleares de aves de corral que son
homólogos a los neutrófilos en los mamíferos. Tienen acción fagocítica primaria
en la respuesta inflamatoria contra agentes infecciosos, como bacterias y hongos
(Rincones & Moreno, 2010). La relación de heterófilos a linfocitos se ha aceptado
como un índice confiable para determinar el estrés en aves de corral. Los
heterófilos son parte de la inmunidad natural y defensa celular contra las
infecciones microbianas, y los linfocitos son células que producen anticuerpos.
Los aumentos en la relación de heterófilos a linfocitos en pollos desafiados se

38
pueden atribuir a una mayor secreción de corticosterona (Rincones & Moreno
Tirado, 2011), lo que finalmente da como resultado una disminución del título de
anticuerpos. En la reciente investigación se muestra una notable diferencia en
cuanto al conteo de heterófilos entre el grupo tratado con la cepa de
Streptomyces sp. RL8 y el control no tratado. Las aves fueron analizadas
minuciosamente post-morten, buscando evidencias de agentes patógenos dentro
del grupo tratado y no se mostraron síntomas infecciosos, lo cual indica que el
agente probiótico ejerció una fuerte influencia sobre el sistema inmune de las
aves. Estos resultados concuerdan con los hallazgos de Sadeghi et al., (2015),
Rincones & Moreno Tirado, (2011) y Gómez, (2013), quienes demostraron que la
suplementación de la dieta con probióticos aumentó significativamente el recuento
de glóbulos blancos en comparación con el grupo de control.
Los valores de eosinófilos aumentaron significativamente en el grupo tratado con
Streptomyces sp. RL8 con relación al grupo control (Tabla 2). Al parecer el
sistema inmune fue estimulado por este microorganismo, efecto que se vio
reflejado con el aumento del bazo como órgano linfoide secundario (Tabla 2),
valores superiores a los encontrados por Sadeghi et al., (2015), Mascheck &
Baker., (2008), donde utilizaron S. cerevisiae, L. acidophilus, B. subtilis y una
mezcla entre ellos.
La concentración de hemoglobina corpuscular media (CHCM) no varió en ninguno
de los dos grupos. El uso de probióticos en forma combinada ha reportado
mejores resultados que al utilizarlos individualmente, lo cual podría explicar la
indiferencia entre los grupos (Bula-Meyer, 1988). Sin embargo, estos valores
difieren de los encontrados por Valderr ama, (2013) quien reportó valores medios
de CHCM en aves alimentadas con probióticos, antibióticos y un grupo control de
225,6; 234,2 y 231,1 g/L, respectivamente. Por el contrario, nuestros resultados
coinciden con Areces & Céspedes, (1992) quienes obtuvieron valores de CHCM
de 331,7 g/L. Es posible según Mascheck & Baker., (2008) que a primera vista, la
diferencia sea grande al confrontarla con nuestros resultados; sin embargo, el tipo
de probiótico así como las condiciones geográficas, climáticas, nutricionales y
sanitarias en que se desarrolló el experimento pudieron influir en el resultado.
Todo parece indicar que la cepa utilizada en esta investigación está ejerciendo
una fuerte influencia sobre el sistema inmune, lo cual pudiera contribuir al
fortalecimiento del mismo.

39
4.3 Conclusiones parciales
Se demuestra que la cepa de Streptomyces sp. RL8 puede ser utilizada como
agente probiótico en pollos de remplazo de la raza Leghorn de 0-21 días de edad,
principalmente como estimulante hematológico y por su función moduladora sobre
el sistema inmune, incrementando los glóbulos blancos en sangre.

40
CAPÍTULO 5. DETERMINACIÓN DE
LAS TRANSFORMACIONES
MICROBIOLÓGICAS EN EL
TRAGTO DIGESTIVO DE LAS AVES
EVALUADAS

41
CAPÍTULO 5. DETERMINACIÓN DE LAS TRANSFORMACIONES
MICROBIOLÓGICAS EN EL TRAGTO DIGESTIVO DE LAS AVES EVALUADAS
Introducción
La alimentación y el manejo de los animales son los factores que más pueden
influir en la manipulación del ecosistema TGI de las aves. Por eso, el empleo de
probióticos es una de las alternativas para contribuir a la estabilidad del
ecosistema GIT e influir en los procesos digestivos y absortivos, lo que determina
el buen funcionamiento del TGI y, por tanto, buen estado de salud del animal
(Collins & Gibson, 1999, Isolauri et al., 2004). Por tal motivo el objetivo de este
capítulo:
1. Determinar las transformaciones microbiológicas en el tracto gastrointestinal de
las aves evaluadas.
5.1 Materiales y Métodos
Se tomaron muestras de los animales evaluados de los intestinos gruesos y
delgados, 1 g por cada segmento, se maceró en un mortero y se le hizo diluciones
hasta 10-7, de los intestinos delgados se sembró las diluciones 10-3 y 10-5 en ACA
(Rangel, 2015), para el conteo de actinomicetos y en Agar McConkey para el
conteo total de E. coli. En el caso de los intestinos gruesos se siembra de forma
similar según García, (2011).
Tratamiento Estadístico
Los datos se procesaron según tratamiento estadístico utilizado en el capítulo III.
En el caso de los conteos de microorganismos, los datos no siguieron una
distribución normal, por lo que se transformaron según logX. Cuando el conteo fue
igual a cero se sumó una constante (X+0,375), según García, (2011). Para todos
los análisis efectuados, el nivel de significación fue de p<0,05.
5.2 Resultados y Discusión
Se encontraron diferencias entre el grupo control y el tratado donde se incluyeron
los microorganismos, lo que indica un estado eubiósico del TGI. Los valores de
los colibacilos se encuentran en el rango de concentraciones normales de la
comunidad microbiana presente en el TGI de los pollos (Mead & Adam, 1975) y
son inferiores a las reportadas por Milián, (2009) y Rondón, (2009) cuando
emplearon preparados probióticos de Lactobacillus y Bacillus, respectivamente,

42
así como los mostrados por García, (2011) al tratar pollos de engorde hasta los
42 días de edad, donde obtuvo un conteo de colibacilos a los 21 días tratados con
L. pentosus LB-31 de 13,44 UFC·g-1 con W. anomalus LV-6 de 9,50 UFC·g-1 y su
mezcla 15,18 UFC·g-1 en pollos de ceba. Olnood et al. (2015) donde usaron cepas
de Lactobacillus johnsonii, Lactobacillus crispatus, Lactobacillus anunidentified
Lactobacillus sp., obtuvieron resultados superiores a los obtenidos en esta
investigación (Olnood et al., 2015). Hoyos et al., (2008) utilizaron EM® (probiótico
comercial), una mezcla de bacterias y levaduras (Lactobacillus cassei
10³UFC/mL, Sacharomyces cerevisiae 10³UFC/mL, Rodhopseudomonas palustris
10³UFC/mL) en concentraciones mayores a 104 UFC/mL de solución en pollos de
engorde. Los resultados del análisis microbiológico mostraron en el día 18 un
recuento de UFC de colibacilos muy por encima a los encontrados en esta
investigación con valores de 82,5 x 10-5 UFC/mL en el lote con EM® y 92.5 x 10-5
UFC/mL en el lote control sin EM®.
El uso de la sepa RL8 y su implementación en el pienso de los pollos demostró
una disminución en el conteo de coliformes totales presentes en el ambiente del
TGI hasta 21 días de edad (Tabla 5). Es importante destacar que una de las
principales vías de contacto y contagio por bacterias patógenas en aves es
precisamente el TGI. El control de las poblaciones de colibacilos es una de las
principales preocupaciones hoy día en la producción avícola tanto a nivel nacional
como internacional, por lo que el uso la cepa de Streptomyces sp. RL8, podría
resultar una alternativa viable para el control de dichos patógenos.

43
Tabla 5: Conteo de actinomicetos y coliformes totales en el tracto gastrointestinal
de los animales evaluados a los 21 días de edad.
Indicadores1 (UFC·g-1 de)
Edad (d)
Tratamientos
Intestino delgado
RL8-H RL8-M RL8-T Cont-H Cont-M Cont-T EE±
Coliformes x10-7
21 6,66 (0,45)a
6,64 (0,44)a
6,65 (0,45)a
0,42 (0)abc
7,25 (1,81)c
6,95 (0,90)ab
0,33
Actinomicetos x10-2
21 2,4 (3,13)a
2,4 (2,91)ab
2,4 (3,02)a
2,4 (2,88)ab
0,42 (0)abc
2,1 (1,44)c
0,82
Intestino Grueso
Edad (d)
RL8-H RL8-M RL8-T Cont-H Cont-M Cont-T EE±
Coliformes x10-9
21 9,08 (1,22)a
9,0 (1,06)a
9,05 (1,14)a
9,32 (2,12)b
9,42 (2,64)c
9,37 (2,38)bc
0,67
Actinomicetos x10-5
21 5,2 (1,96)a
5,3 (2,24)ab
5,3 (2,10)ab
4,7 (0,50)c
4,6 (0,43)c
4,6 (0,47)c
0,62
a,b,c, Por columna las medias con diferentes letras difieren a p<0,05 (Duncan, 1955). 1Datos
transformados según logX, ( ) Datos originales. RL8-H: Media de las UFC·g-1 tratados con la cepa RL8
en pollos Hembras, RL8-M: Media de las UFC·g-1 tratados con la cepa RL8 en pollos Machos, RL8-T:
Media Total de las UFC·g-1 tratados con la cepa RL8, Cont-H: Media de las UFC·g-1 del grupo control
Hembra sin probiótico, Cont-M: Media de las UFC·g-1 del grupo Control Macho sin probiótico, Control-
T: Media total de las UFC·g-1 del grupo control sin probiótico, EE±: Error estándar de la media.
5.3 Conclusiones parciales
La cepa Streptomyces sp. RL8 ejerce control sobre las poblaciones de colibacilos
presentes en el tracto gastrointestinal de las aves evaluadas hasta los 21 días de
edad.

44
CONCLUSIONES GENERALES

45
CONCLUSIONES GENERALES
Se demostró que la cepa de Streptomyces sp. RL8 es capaz de tolerar y sobrevivir a
altas concentraciones de sales biliares y cloruro de sodio, además de resistir
bajos valores de pH.
Al evaluar los efectos de la cepa sobre los parámetros bioproductivos, de salud y
morfométricos de las aves, de determinó que esta no influye significativamente
sobre el peso de los animales, los órganos evaluados mostraron una moderada
diferencia, aunque se evidenció que puede inducir cambios en la morfometría de
los intestinos.
La cepa de Streptomyces sp. RL8 mejoró los parámetros hematológicos e
inmunológicos en pollos de la raza Leghorn.
Se evidenció una gradual disminución en los conteos de las poblaciones de
colibacilos los intestinos delgados y gruesos, lo cual acredita a la cepa de
Streptomyces sp. RL8 como un buen agente controlador de colibacilos en estos
segmentos digestivos.
Los resultados obtenidos in vitro e in vivo, demuestran que la cepa de
Streptomyces sp. RL8 posee propiedades probióticas y puede ser utilizada como
aditivo microbiano destinado a la alimentación de pollos recién eclosionados para
controlar la colibacilosis, fortalecer su microbiota intestinal, reduciendo las
poblaciones de colibacilos a nivel del TGI, promover altos valores hematológicos,
estimular su sistema inmune mediante una buena producción de heterófilos y
linfocitos, controlar el crecimiento de patógenos oportunistas para si, incrementar
los índices bioproductivos del sector avícola.

46
RECOMENDACIONES

47
RECOMENDACIONES
Realizar estudios de antagonismo in vitro con cepas de E. coli patógenas para
pollos.
Hacer combinaciones con otros posibles agentes probióticos que se utilizan en la
avicultura como Lactobacillus ssp., Bacillus ssp., levaduras, etc.
Aumentar el tamaño de muestra de las aves evaluadas.
Realizar pruebas de hemaquimica detallada (glicemia, colesterol, proteinas
totales, etc).
Realizar pruebas inmunológicas más específicas, como por ejemplo: títulos de
anticuerpos ante determinadas vacunas, y frente a la cepa RL8, conteos de
linfocitos T y B, etc.
Continuar el estudio a otras fases de crecimiento en gallinas Leghorn, así como
su evaluación, caracterización y efecto hasta la puesta del huevo.
Trabajar fundamentalmete con pollos de engorde raza Broiler (productores por
excelencia de carne).
Establecer una relación beneficio-costo, análisis económico del uso de la cepa
RL8 en la producción de pollos de Leghorn en el sector avícola.

48
REFERENCIAS BIBLIOGRÁFICAS

49
REFERENCIAS BIBLIOGRÁFICAS
ANGEL, R., WOO KIM, S., LI, W. & JIMENEZ-MORENO, E. 2013. Velocidad de
paso y pH intestinal en aves: implicaciones para la digestión y el uso de enzimas.
Departament of Animal and Avian Sciences. University of Meryland College Park,
MD 20742, USA
ANGELAKIS, E. 2017. Weight gain by gut microbiota manipulation in productive
animals. Microbial Pathogenesis 106 162-170.
ARECES, A. J. & CÉSPEDES, N. 1992. Potencialidad productiva de algunas
caragenófitas del Indopacífico en aguas del Caribe. Boletín de la Red
Latinoamericana de Acuicultura, 6 (2), 13-16.
ASGHAR SADEGHI, A., SHAWRANG, P. & SHAKORZADEH, S., . 2015. Immune
Response of Salmonella Challenged Broiler Chickens Fed Diets Containing
Gallipro®, a Bacillus subtilis Probiotic. Probiotics & Antimicro.
BAGHLAF, A. O., ABOU-ZEID, A. A., I. EL-DIWANY, A., EL-WAHAB, A., EISSA,
I., FOUAD, M. & YESSEIN, M. 1979. Production of oxytetracycline, by
streptomyces rimosus 12907, as an animal feed supplement. Agricultural Wastes,
1, 301-310.
BERNAL, M. G., CAMPA-CÓRDOVA, Á. I., SAUCEDO, P. E., GONZÁLEZ, M. C.,
MARRERO, R. M. & MAZÓN-SUÁSTEGUI, J. M. 2015. Isolation and in vitro
selection of actinomycetes strains as potential probiotics for aquaculture.
Veterinary World, 8, 170-176.
BERNAL, M. R. G. 2016. OBTENCIÓN DE ACTINOMICETOS MARINOS CON
ACCIÓN PROBIÓTICA EN OSTIONES Y CAMARONES.
BEZKOROVAINY, A. 2001. Probiotics: Determinants of survival and growth in the
gut. Am. J. Clin. Nutr, 73, 399S-405S.
BRADFORD, M. 2000. Algas: las verduras del mar. Los nutritivos tesoros para la
salud y el paladar. Océano, Barcelona. España, 191.

50
BULA-MEYER, G. 1988. Cultivos y utilización comercial de las macroalgas
marinas. Colombia. , Universidad Tecnológica de Magdalena, Facultad de
Ingeniería Pesquera, Serie de Manuales Técnicos, Colombia, 57.
BYRD, J., ANDERSON, R., R CALLAWAY, T., W MOORE, R., D KNAPE, K., F
KUBENA, L., L ZIPRIN, R. & NISBET, D. 2003. Effect of experimental chlorate
product administration in the drinking water on Salmonella typhimurium
contamination of broilers.
CASTANON, J. I. R. 2007. History of the use of antibiotic as growth promoters in
European poultry feeds. Poultry Science, 86, 2466–2471.
COLLINS, M. D. & GIBSON, G. R. 1999. Probiotics, prebiotics, and synbiotics:
approaches for modulating the microbial ecology of the gut. Am. J. Clin. Nutr, 69
(1), 1042-1057.
CONTRERAS, M. & FERNÁNDEZ, J. F. 1999. Entidades inmunosupresoras y su
correlación com otras condiciones patológicas y de la crianza. Industria Avícola,
29-34.
CROSBY, W. H., MUNN, J. I. & FURTH, F. W. 1954. Standardizing a method for
clinical hemoglobinometry. U.S. Armed Forces Med, 693-703.
CUETO-VIGIL, M. C., ACUÑA-MONSALVE, Y. & VALENZUELA-RIAÑO, J. 2010.
Evaluación in vitro del potencial pobiótico de bacterias ácido lácticas aisladas de
suero costeño. Actual Bio,l 32 (93, 129-138.
CHICHLOWSKI, M., CROOM, J., MCBRIDE, B. W., HAVENSTEIN, G. B. & KOCI,
M. D. 2007. Metabolic and physiological impact of probiotics or direct-fed-
microbials on poultry: A brief review of current knowledge. Int. J. Poult. Sci,6, 694-
704.
DANFORTH, H. D., RUFF, M. D., REID, W. M. & MILLER, R. L. 1977.
Anticoccidial Activity of Salinomycin in Battery Raised Broiler Chickens. Poultry
Science, 56, 926-932.

51
DEIVER HOYOS, H., NELSON ALVIS, G., LEONEL JABIB, R., MARINA
GARCÉS, B., DALIS PÉREZ, F. & SALIM MATTAR, V. 2008. Utility of effective
microorganisms em® in an avian farm of cordoba: Productives parameters and
enviromental control. Rev.MVZ Córdoba, 13 (2), 1369-1379.
EHRMANN, M. A., KURZAK, P., BAUER, J. & VOGEL, R. F. 2002.
Characterization of lactobacilli towards their use as probiotic adjuncts in poultry.
Journal of Applied Microbiology, 92, 966-97.
FAO 2016. Estado mundial de la pesca y la acuicultura. Organización de las
Naciones Unidas para la Alimentación y la Agricultura. Disponible en:
http://www.fao.org/3/a-i5555s.pdf (revisada febrero 2017).
FAO/WHO 2002. (Food and Agriculture Organization of the United Nations/World
Health Organization). Guidelines for the evaluation of probiotics in food. Report of
a joint FAO/WHO working group on drafting guidelines for the evaluation of
probiotics in food. April 30 and May 1. London Ontario, Canada. Disponible en:
http://www.who.int/foodsafety/fs_management/en/probiotic_guidelines.pdf FAO
2012. Global Aquaculture Production. Organización de las Naciones Unidas para
la Alimentación y la Agricultura. Roma.
FARFÁN-GARCÍA A .E., ARIZA-ROJAS S.C., VARGAS-CÁRDENASF. A. & V.,
V.-R. L. 2016. Mecanismos de virulencia de Escherichia coli enteropatógena.
Patogenia.
FRIZZO, L., SOTO, L., BERTOZZI, E., SEQUIERA, E., MARTI, L. & ROSMINI, M.
2006. Evaluación in vitro de las capacidades probióticas orientadas al diseño de
inóculos probióticos multiespecie para ser utilizados en la crianza de terneros.
Rev FAVE, 5, 1-2.
GADDE, U. D., OH, S., LILLEHOJ, H. S. & LILLEHOJ, E. P. 2018. Antibiotic
growth promoters virginiamycin and bacitracin methylene disalicylate alter the
chicken intestinal metabolome. Scientific Reports, 8, 3592.

52
GARCÍA, H. Y. 2011. Obtención de Microorganismos con Actividad Probiótica a
Partir de Excretas de Pollos de Ceba Fermentadas.
GATESOUPE, F. J. 1999. The use of probiotics in aquaculture. Aquaculture, 180,
147-165.
GEORGE, B. A., FAGERBERG, D. J., QUARLES, C. L., FENTON, J. M. &
MCKINLEY, G. A. 1982a. Effect of bambermycins on quantity, prevalence,
duration, and antimicrobial resistance of Salmonella typhimurium in experimentally
infected broiler chickens. American Journal of Veterinary Research, 43(2), 299-
303.
GEORGE, B. A., QUARLES, C. L. & FAGERBERG, D. J. 1982b. Virginiamycin
Effects on Controlling Necrotic Enteritis Infection in Chickens. Poultry Science, 61,
447-450.
GILDBERG, A., MIKKELSEN, H., SANDAKER, E. & RINGØ, E. 1997. Probiotic
effect of lactic acid bacteria in the feed on growth and survival of fry of Atlantic cod
(Gadus morhua). In Asia-Pacific Conference on Science and Management of
Coastal Environment. Springer, 279-285.
GIRAFFA, G., CHANISHVILI, N. & WIDYASTUTI, Y. 2010. Importance of
lactobacilli in food and feed biotechnology. Res. Microbiol, 161, 480-487.
GOETTING, V., LEE, K. A. & TELL, L. A. 2011. Pharmacokinetics of veterinary
drugs in laying hens and residues in eggs: a review of the literature. Veterinary
Phamacology and Therapeutic, 10 (1111), 1365-2885.
GÓMEZ, E. 2013. Evaluación nutricional y propiedades biológicas de algas
marinas comestibles. Estudios in vitro e in vivo. Trab. Grad. Dr. Farmacia y
Tecnología Farmacéutica. Universidad Complutense de Madrid, Madrid, España,
218.

53
GORNATTI-CHURRIA, C. D., CRISPO M., SHIVAPRASAD H. L. & A., U. F. 2018.
Gangrenous dermatitis in chickens and turkeys. Journal of Veterinary Diagnostic
Investigation, 30 (2), 188.
GUPTA, V. K., TUOHY, M. G., LOHANI, M. & O’DONOVAN, A. 2015.
Antimicrobials from medicinal plants: Research initiatives, challenges, and the
future prospects.
IRATXE PÉREZ, A. 2015. Calidad y seguridad microbiológica de la sangre de
pollo como especial referencia a la incidencia de Salmonella, Campylobacter y
Listeria monocytogenes en las distintas etapas de la producción de procesado.
25-40.
IRIANTO, A. & AUSTIN, B. 2002. Probiotics in aquaculture. J. Fish. Dis, 25, 633-
642.
ISOLAURI, E., SALMINEN, S. & OUWEHAND, A. C. 2004. Probiotics. Best
Practice. Res. Clin. Gastroenterol, 18, 299-313.
KAZUE , L., BIONDARO GÓIS, M., REGINA DE MORAES GARCIA, E. & ESPINO
HERNÁNDEZ, M. 2015. Variations on the Efficacy of Probiotics in Poultry.
KENNEDY, D. G., CANNAVAN, A. & MCCRACKEN, R. J. 2000. Regulatory
problems caused by contamination, a frequently overlooked cause of veterinary
drug residues. Journal of Chromatography A, 882, 37–52.
KESARCODI-WATSON, A., KASPAR, H., LATEGAN, M. J. & GIBSON, L. 2008.
Probiotics in aquaculture: the need, principles and mechanisms of action and
screening processes. Aquaculture, 274, 1-14.
LANNOT 2005. Bergey's Manual of Systematic Bacteriology: Volume 3: The
Firmicutes.
LARA-FLORES, M. 2011. The use of probiotic in aquaculture: an overview.
International Res. J. Microbiol, 2, 471-478.

54
LINO-NAVARRO, M., LEÓN QUISPE, J. & HUAMÁN ITURRIZAGA, M. 2016.
Evaluación de la capacidad antagonista de un antimicrobiano producido por
Streptomyce sp. CEPA 13A-2 frente a microorganismos resistentes a B-
Lactámicos de origen hospitalario. Rev Peru Investig Matern Perinat, 5(1), 28-34.
MASCHECK, J. & BAKER., R. 2008. Macroalgal chemical defenses and their roles
in structuring tropical marine communities. Algal Chemical Ecology. C. Asmsler
(ed.). Editorial Springer, Berlin, Alemania, 313.
MATEOS, G. G., LÁZARO, R. & GRACIA, M. I. 2002. Modificaciones nutricionales
y problemática digestiva en aves. XVIII Curso de especialización FEDNA. 4-5 de
noviembre. Madrid, España, 15-37.
MCHUGH, D. 2002. Perspectivas para la Producción de Algas Marinas en los
Países en Desarrollo. FAO Circular de Pesca No. 968, Disponible en:
http://www.fao.org/docrep/004/y3550s/y3550s04.htm (revisada diciembre 2015).
MEAD, G. C. & ADAM, B. W. 1975. Some observations on the caecal microflora of
the chick during the first two weeks of life. 169-176.
MEHDI, Y., MARIE-PIERRE, L., GAUCHER, M-L., CHRFI, Y., SURESH, G.,
ROUISSI, T., KAUR BRA, S., AVALOS RAMIREZ, A. & GODBOUT, S. 2018. Use
of antibiotics in broiler production: Global impacts and alternatives. Animal
Nutrition, 4,170-178.
MILIÁN, G. 2009. Obtención de cultivos de Bacillus spp. y sus endosporas.
Evaluación de sus actividad probiótica en pollos (Gallus gallus domesticus).
Instituto de Ciencia Animal. La Habana, Cuba, 134.
NEMUTANZHELA, M. E., ROETS, Y., GARDINER, N. & LALLO, R. 2014. The use
and benefits of Bacillus based biological agents in aquaculture. En: Hernandez-
Vergara, M. (ed.) Sustainable aquaculture techniques. Murcia, Spain.
NEWAJ-FYZUL, A., AL-HARBI, A. H. & AUSTIN, B. 2014. Developments in the
use of probiotics for disease control in aquaculture. Aquaculture, 431, 1-11.

55
OLNOOD, C. G., SLEMAN, S. M., MINGAN, C. & PAUL, A. 2015. Novel
probiotics:Their effect son growth performance, gut development, microbial
community andactivity of broiler chickens. Animal Nutrition, 1, 184–191.
ONGOR, H., KALIN, R., KARAHAN, M., CETINKAYA, B., MCAULIFFE, L. &
NICHOLAS, R. A. J. 2008. Isolation of Mycoplasma bovis from broiler chickens in
Turkey. Avian Pathology, 37, 587-588.
PARK, S., HWANG, Y., KIM, Y., KIM, J., SONG, J., LEE, K., JEONG, K., RHEE,
M., KIM, K. & KIM, T. 2006. Comparison of pH and bile resistance of Lactobacillus
acidophilus strains isolated from rat, pig, chicken, and human sources. Journal of
Microbiology and Biotechnology, 22 (1), 35-37.
PENSACK, J. M., WANG, G. T. & SIMKINS, K. L. 1982. Avoparcin — A Growth-
Promoting Feed Antibiotic for Broiler Chickens. Poultry Science, 61, 1009-1012.
PÉREZ. M., M. LAURENCIO. M., RONDÓN. A. J., MILIAN. G., BOCOURT. R. &
F., A. 2011. Actividad Antimicrobiana de una mezcla probiótica de exclusión
competitiva y su estabilidad en el tiempo. Salud Anim, 33, 147-153.
PEROZO-MARÍN, F., NAVA, J., MAVÁREZ, Y., ARENAS, E., SERJE, P. &
BRICEÑO, M. 2004. Caracterización morfométrica de los órganos linfoides en
pollos de engorde de la línea Ross criados bajo condiciones de campo en el
Estado Zulia, Venezuela. Revista Científica, FCV-LUZ, 14, 217-225.
PROCÓPIO, R.E., MARTINS, M.K., AZEVEDO, J.L., ARAÚJO, J.M. 2012.
Antibiotics produced by Streptomyces.
PUIG PEÑA, M.Y., ESPINO HERNÁNDEZ & CASTILLO, V. L. 2011. Resistencia
antimicrobiana en Salmonella y E. coli aisladas de alimentos: revisión de la
literatura. Panorama Cuba y Salud 2011.
RAHIMI, S.H. & KHAKSEFIDI, A. 2006. A comparison between the effects of a
probiotic (Bioplus 2B) and an antibiotic (virginiamycin) on the performance of

56
broiler chickens under heat stress condition. Iranian Journal of Veterinary
Research, University of Shiraz, 7(3), 16.
RANGEL, M. G. B. 2015. Plantas Toxicas para el Ganado en el Noreste de
Coahuila. .
RINCONES, R. E. & MORENO, D. 2010. Proyecto desarrollo experimental de la
maricultura de algas como alternativa productiva a las comunidades costeras
entre los corregimientos de Carrizal y Cabo de La Vela, departamento de La
Guajira Informe Final. Convenio Fundación Terrazul-SENA (Manuscrito no
publicado).
RINCONES, R. E. & MORENO TIRADO, D. A. 2011. Technical and economical
aspects for the commercial establishment of seaweed mariculture in Colombia:
Experiences in the Guajira Peninsula. Bogotá (Colombia), Volumen XV (28).
ROCHA, A. K. M. 2012. Uso de Antimicrobianos en la Avicultura: Sus
Implicaciones en la Salud Pública.
RONDÓN, A. J. 2009. Obtención de biopreparados a partir de lactobacilos
autóctonos del tracto digestivo de pollos y evaluación de su efecto probiótico en
estos animales. Instituto de Ciencia Animal. La Habana, Cuba, 131.
SOKAL, R.R & ROHLF, F. 1995. Biometry (3d edn). New York: WH Feeman and
Company. p. 98.
SCHREZENMEIR, J. & DE VRESE, M. 2001. Probiotics, prebiotics, and synbiotics
approaching a definition. J. Clin. Nutr, 73, 361-364.
SOLECKA, J., ZIEMSKA, J., POSTEK, M. & RAJNISZ-MATEUSIAK, A. 2012.
Biologically active secondary metabolites from Actinomycetes.
TAHERI, H. R., MORAVEJ, H., TABANDEH, F., ZAGHARI, M. & SHIVAZAD, M.
2009. Screening of lactic acid bacteria toward their selection as a source of
chicken probiotic. Poultry Science, 88, 1586-1593.

57
TANAKA, H., KOYAMA, Y., AWAYA, J., MARUMO, H., OIWA, R., KATAGIRII, M.,
NAGAI, T. & S., O. 1975. Nanomycins, new antibiotics produced by a strain ok
Streptomyces. The Journal of Antibiotic, 28 (11), 860-867
TINH, N. T. N., LINH, N. D., WOOD, T. K., DIERCKENS, K., SORGELOOS, P. &
BOSSIER, P. 2007. Interference with the quorum sensing systems in a Vibrio
harveyi strain alters the growth rate of gnotobiotically cultured rotifer Brachionus
plicatilis. J. Appl. Microbiol, 103, 194-203.
TUOMOLA, E. M., CRITTENDEN, R., PLAYNE, M. & ISOLAURI, E., ALMINEN, S.
J. 2001. Quality assurance criteria for probiotic bacteria. American Journal of
Clinical Nutrition, 73, 393-398.
USDA‐FAS 2018. Livestock and Poultry: World Markets and Trade.
USDA‐FAS 2016. Livestock and Poultry: World Markets and Trade.
VALDERR AMA, D. 2013. Social and economic dimensions of carrageenan
seaweed farming. FAO Technical Paper, Roma., 580.
VERSCHUERE, L., HEANG, H., CRIEL, G., SORGELOOS, P. & VERSTRAETE,
W. 2000. Selected bacterial strains protect Artemia spp. from the pathogenic
effects of Vibrio proteolyticus CW8T2. Appl. Environ. Microbiol, 66, 1139-1146.
VINE, N. G., LEUKES, W. D. & KAISER, H. 2006. Probiotics in marine larviculture.
FEMS Microbiol. Rev, 30, 404-427.
VOIGT, G. L. 2003. Conceptos y técnicas hematológicas para técnicos
veterinarios. Zaragoza, ACRIBIA, 144.
WHITTOW, G. C. 2000. Sturkie’s Avian Physiology. Sturkie’s Avian Physiology.
Academic Press, London, 5.
WHO. 2002. Guidelines for the evaluation of probiotics in food. Report of a joint
FAO/WHO working group on drafting guidelines for the evaluation of probiotics in

58
food. Food and Agriculture Organization. Fecha de acceso: 17 de diciembre de
2010. Disponible en:
<http://www.who.int/foodsafety/fs_management/en/probiotic_guidelines.pdf>.
XU, S., LIN, Y., ZENG, D., ZHOU, M., ZENG, Y., WANG, H., ZHOU, Y., ZHU, H.,
PAN, K., JING, B. & NI, X. 2018. Bacillus licheniformis normalize the ileum
microbiota of chickens infected with necrotic enteritis. Scientific Reports, 8, 1744.

59
ANEXOS

60
ANEXOS
Anexo 1. Composición de la dieta para pollos de Leghorn al inicio de crecimiento.
Ingredientes, % en base seca Inicio (0-21 d)
Harina de maíz 45,60
Harina de soya (42 % proteína bruta) 43,86
Aceite vegetal 5,39
Premezcla minero-vitamínica* 1,00
Cloruro de sodio 0,37
Fosfato monocálcico 1,80
Carbonato de calcio 1,60
DL-metionina 0,28
Lisina 0,10
Anexo 2. Medios de cultivos empleados
CALDO TRIPTICASA SOYA (TSB)
Componentes g/l
Tripticasa 17
Peptona 3
Cloruro de sodio 5
Fosfato dipotásico 2,5
Dextrosa 2,50
Agua destilada 1L
AGAR CASEÍNA–ALMIDÓN
Componentes g/L
Almidón 10,0
Caseína 0,30
Nitrato de Potasio 2,0
NaCl 2,0
K2HPO4 2,00
MgSO4.7H2O 0,05
CaCO3 0,02
FeSO4.7H2O 0,01
Agar 15,0
Agua destilada 1000 mL
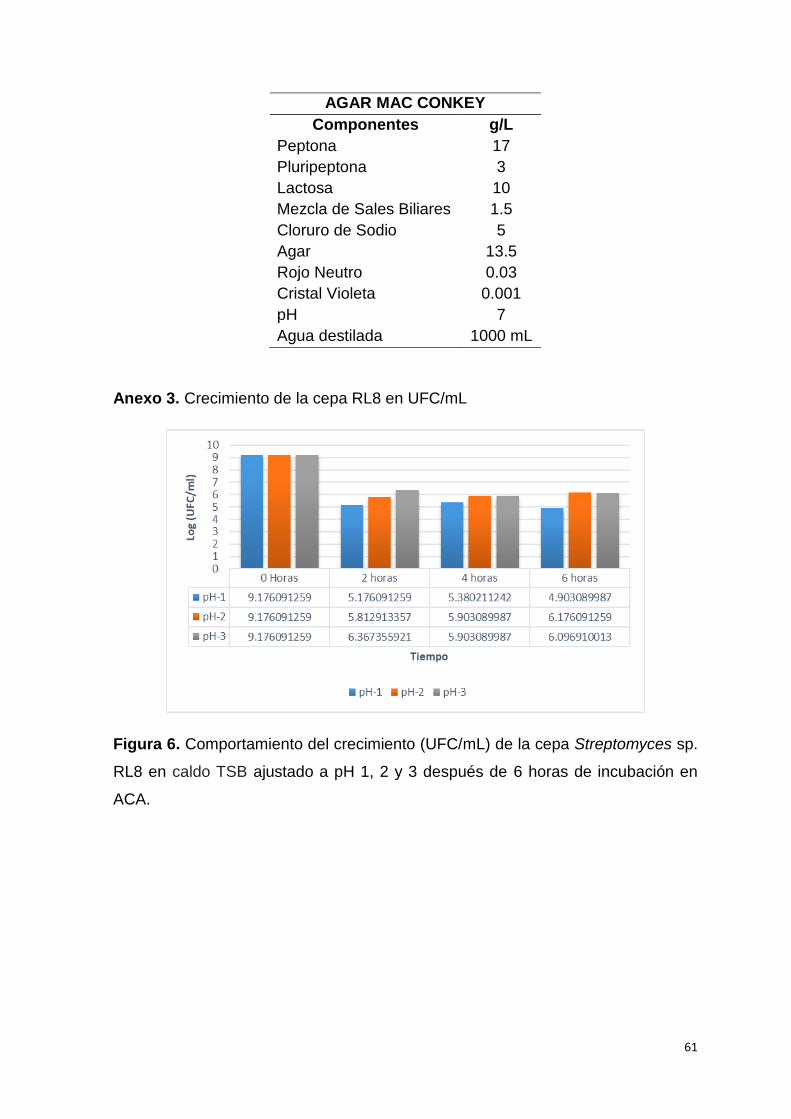
61
AGAR MAC CONKEY
Componentes g/L
Peptona 17
Pluripeptona 3
Lactosa 10
Mezcla de Sales Biliares 1.5
Cloruro de Sodio 5
Agar 13.5
Rojo Neutro 0.03
Cristal Violeta 0.001
pH 7
Agua destilada 1000 mL
Anexo 3. Crecimiento de la cepa RL8 en UFC/mL
Figura 6. Comportamiento del crecimiento (UFC/mL) de la cepa Streptomyces sp.
RL8 en caldo TSB ajustado a pH 1, 2 y 3 después de 6 horas de incubación en
ACA.