1. los biologos en el tiempoecaths1.s3.amazonaws.com/catbioveg/1898664168.notas... · establecer...
TRANSCRIPT

1 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
1. LOS
BIOLOGOS EN EL
TIEMPO Este ensayo trata de describir los principales
cambios que han sufrido los biólogos
vegetales y el conocimiento biológico en
relación con los tipos de sociedades que
hacían uso de él. En este sentido se pueden
establecer una serie de etapas históricas. Las
fechas que se dan para estas etapas son
aproximadas y en ningún caso los cambios son
abruptos y tampoco implican la finalización
inmediata de las etapas previas.
Los biólogos cazadores-recolectores, activos
desde el comienzo de los tiempos…
Los humanos primitivos, cazadores-
recolectores (¿o recolectores-cazadores?),
poseían un conocimiento amplio y detallado
de un número grande de especies vegetales y
animales, usualmente en el orden de varios
cientos y hasta miles de especies. Ese
conocimiento permitía clasificar a las plantas
de acuerdo a su utilidad o función:
comestibles, tóxicas, medicinales, industriales,
etc.
En la actualidad todavía existen comunidades
de cazadores-recolectores que mantienen este
tipo de cultura biológica. Entre estos ejemplos
modernos están los bosquimanos de las
regiones desérticas de Sudáfrica, algunos
aborígenes australianos y algunas tribus
indígenas del Amazonia. Estamos asistiendo a
su desaparición que probablemente tendrá
lugar en las próximas décadas.
En estas comunidades todos los miembros
tienen en general un gran conocimiento
biológico, pueden identificar numerosas
especies (cientos o miles) y conocen sus
propiedades. Además del gran conocimiento
biológico generalizado en estas sociedades,
existen individuos especialistas: shamanes o
brujos, frecuentemente con un enfoque
religioso y médico combinado, quienes tienen
conocimiento superiores al promedio de la
comunidad.
Aparentemente entre los 10.000 y 20.000 AP,
aparecieron los proto-agricultores, que sin
cultivar, “favorecían” determinadas especies
(ej,eliminando competencia, usando el fuego)
y planeaban sus migraciones estacionales en
función de la fenología de estas.
10.000 (30.000-7.000) años AP: Comienzan
los biólogos agricultores
Un cambio trascendental se produjo
aproximadamente hace 10.000 años en Oriente
Medio, y más tarde se difundió en el resto de
Asia, Europa y África. Este cambio consistió
en la domesticación de los cultivos,
principalmente cereales. Alrededor de 8.000
años atrás un cambio similar se produjo en
América con la domesticación del maíz y una
serie de tubérculos.
El desarrollo de la agricultura fue
probablemente el cambio tecnológico más
importante para la humanidad, ya que permitió
obtener excedentes alimenticios que con el
tiempo indujeron los profundos cambios
sociales y políticos que llevaron al desarrollo
de la cultura moderna. Esta puede sintetizarse
como un proceso de especialización cultural
progresiva en algunos miembros de la
sociedad y una transformación profunda del
paisaje para adaptarlo a la agricultura.
Este cambio, llamado la revolución agrícola,
se produjo después de la domesticación de
distintas especies de animales. Sin embargo
tuvo consecuencias políticas y sociales mucho
más profundas ésta.
En la actualidad, los biólogos agricultores
organizados de distintas formas (campesinos,
productores agropecuarios, ingenieros
agrónomos, multinacionales de agro-
alimentos, etc.) son los responsables de
alimentar a gran parte de la humanidad. La
mayor parte de las tierras planas más
productivas del mundo han sido transformadas
con ese fin. En las sociedades campesinas los

2 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
agricultores representaban el 90% o más de la
población. En las sociedades modernas en
cambio están reducidos al 5%, o menos aún.
500 AC-1600 DC: Los primeros biólogos
científicos.
La división de roles en las sociedades
agrícolas permitió la aparición de individuos
especializados en razonar y acumular
conocimientos: los científicos. Un poco
arbitrariamente puede ponerse el comienzo de
esta etapa en la Grecia clásica, donde los
grandes filósofos concentraron la suma del
conocimiento contemporáneo, incluyendo las
ciencias naturales. Sin embargo, la
información biológica fue manejada en mayor
cantidad y calidad por los herederos de los
shamanes o brujos: los médicos y los
alquimistas. Durante muchos siglos estos
profesionales concentraron el mayor esfuerzo
al conocimiento vegetal, apuntando
principalmente a fines terapéuticos.
En muchas culturas estos profesionales se han
mantenido hasta nuestros días. Por ejemplo en
la cultura andina existen algunos grupos
étnicos que se han especializado en esto: son
los Calawayas o los Yungas, que hasta no hace
mucho todavía entraban en territorio argentino
desde Bolivia.
Durante la Edad Media el conocimiento
científico y la herencia académica griega
quedó confinada a los monasterios, que
también cumplieron un rol en el desarrollo de
la biología.
Los médicos, alquimistas, curanderos y
religiosos fueron los encargados de la
conservación y el desarrollo de las ciencias
biológicas y la biología vegetal en particular
durante muchos siglos. El médico belga Van
Helmont fue por ejemplo el primero en dar
una pista sobre las bases de la fotosíntesis,
alrededor de 1600. Sus observaciones fueron
complementadas más tarde (1700-1800) por el
clérigo inglés Priestley y el médico holandés
Ingenhousz. Los experimentos y
razonamientos de estos científicos permitieron
entender el proceso de fotosíntesis tal como lo
aceptamos actualmente.
Siglo XVIII-presente: los biólogos estatales
Hasta el siglo 18 el conocimiento biológico
estuvo en manos de profesionales que usaban
la biología más como herramienta que como
objetivo, es decir eran médicos, curanderos, o
afines, o bien tenían su sustento asegurado de
otra manera: eran religiosos o de la nobleza.
Con la revolución industrial y el surgimiento
de las potencias coloniales europeas tomó gran
importancia un nuevo tipo de profesional de la
biología: el biólogo estatal.
El desarrollo de la nomenclatura científica
tuvo lugar entre los siglos XIII y XVIII, hasta
la publicación del Species Plantarum, por Carl
von Linné en 1753. Linné o Lineo fue un
naturalista y académico sueco que desarrolló
las bases del sistema moderno de clasificación
de los seres vivos.
Una de las primeras investigaciones de los
biólogos estatales fue describir la biota de
países extraños y la obtención de información
potencialmente útil para distintos fines. Tal
vez el biólogo más destacado en este rol fue
Charles Darwin, quién viajó como
acompañante en un viaje financiado por la
corona inglesa alrededor del mundo durante
cinco años (1831-1836). Ese viaje le permitió
realizar observaciones y recopilar información
que resultaron esenciales para desarrollar la
teoría de la evolución.
En ese período surgieron los primeros
herbarios y jardines botánicos modernos
(Kew, París, Berlín, San Petersburgo). Con el
desarrollo de los estados modernos surgieron
las universidades modernas y posteriormente
los centros de investigación, con un
crecimiento exponencial del número de
biólogos estatales. De este modo la biología se
afianzó como una ciencia independiente.
Mediados siglo XX: los “biólogos
controladores” y “biólogos privados”.
En el siglo XX, particularmente en la segunda
mitad han comenzado a aparecer variantes de

3 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
biólogos científicos. Algunos de ellos venden
sus servicios profesionales a grandes empresas
o a organizaciones no gubernamentales
(ONGs: UICN, WWF, FVSA, Greenpeace) en
el campo de la gestión de medio ambiente y la
biodiversidad (“biologos privados”).
Por otro lado, también en este período el
estado comenzó a enrolar biólogos para el
control de los problemas medio ambientales y
para la protección de distintas áreas naturales.
Surgieron una serie de especializaciones de
biólogos en los organismos de control estatal,
como Parques Nacionales, Provinciales, etc.
(“biólogos controladores”).
Actualmente también hay un número creciente
de biólogos dedicados a la enseñanza de la
biología. Mientras que el comienzo de la
humanidad, todos los hombres eran biólogos,
actualmente lo son apenas una pequeña
fracción. Por ello el rol de quienes ense ñan
biología es particularmente trascendente y de
gran impacto potencial en la conducta de la
sociedad.
Figuras 1.1. A y B. Miguel Lillo (1862.1931) fue
probablemente el biólogo más destacado de los
siglos XIX y XX en Tucumán. Fue empleado
estatal como profesor de química, pero también
con una fortuna personal que le permitió realizar
importantes viajes, colecciones de plantas y
desplegar una intensa actividad científica. Aquí en
una fotografía de la época y un dibujo realizado
por el mismo.

4 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
2.CELULA
VEGETAL
Generalidades
Las células son las unidades estructurales y
funcionales de la vida. Los organismos más
pequeños están constituídos por una sola célula,
mientras que los más grandes poseen algunos
miles de millones de células. Existen numerosos
tipos diferentes de células. Por otro lado, a la par
de esta gran diversidad, las células tienen una
notable similitud.
La teoría celular, cuyo enunciado es simplemente
que todos los organismos están constituídos por
células, junto con la teoría de la evolución, son dos
grandes conceptos unificadores de las ciencias
biológicas desarrollados en el siglo pasado.
Cada célula es una unidad, auto-contenida, y
autosuficiente al menos en parte, encerrada en una
membrana (membrana plasmática = membrana
celular = plasmalemma) que controla el pasaje de
sustancias hacia y desde el interior, haciendo
posible que la célula difiera quimicamente y
estructuralmente del entorno. Encerrado por esta
membrana se encuentra el citoplasma, que en la
mayor parte de las células incluye una serie de
corpúsculos y sustancias disueltas o en suspensión.
Entre éstos componentes, cada célula posee ADN,
(ácido desoxiribonucleico), que contiene la
información genética necesaria para el
funcionamiento y reproducción.
Procariontes y eucariontes
Procariontes son organismos unicelulares que
carecen de núcleo diferenciado, es decir una
envoltura membranosa que rodea el ADN célular.
El ADN tampoco se encuentra asociado a
proteínas formando estructuras cromosómicas.
En los eucariontes el ADN se encuentra asociado a
proteínas formando cromosomas, encerrados por
una doble membrana formando un núcleo. Además
en las células eucarióticas existen otras estructuras
rodeadas por membranas, denominados orgánulos
celulares.
La célula vegetal
Las células vegetales típicas están constituidas por
una pared celular más o menos rígida y un
protoplasma. El protoplasma está constituido por
el citoplasma y el núcleo. El citoplasma incluye
una serie de entidades independientes separadas
por membranas (orgánulos u organelas), distintos
sistemas de membranas y entidades no
membranosas. Todas estas estructuras están
suspendidas en una "sopa celular", el citosol o
matriz celular. El citosol está separado de la pared
celular por una membrana celular. El citoplasma se
encuentra frecuentemente en movimiento. Este
movimiento es denominado ciclosis, que suele
mantenerse en tanto la célula se encuentre con vida

5 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
. Figura 2.1. Esquema de una célula vegetal con sus diferentes orgánulos y estructuras celulares.

6 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Inventario de componentes de una célula vegetal
A. Pared celular Laminilla media
Pared primaria
Pared secundaria
Plasmodesmas
B. Protoplasma Núcleo Membrana nuclear
Nucleoplasma
Cromatina
Nucleolo
Citoplasma Citosol
Orgánulos con doble membrana
Mitocondrias
Plastidios
Orgánulos con una membrana
Microcuerpos
Vacuolas
Sistema de endomembranas
Retículo endoplasmático
Dictiosomas
Vesículas
Membrana plasmática
Citoesqueleto
Microtúbulos
Microfilamentos
Ribosomas
C. Flagelos
D. Sustancias ergásticas (pueden presentarse en la pared o el protoplasma)
Cristales
Antocianinas
Gránulos de almidón
Taninos
Grasas, aceites, ceras
Cuerpos protéicos

7 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Membrana plasmática
La membrana plasmática tiene una típica
estructura de tres capas (modelo de Mosaico
Fluido), visible al microscopio electrónico. Está
compuesta predominantemente de lípidos y
proteínas. Posee una serie de funciones muy
importantes:
media en el transporte de sustancia hacia y
desde el interior de la célula.
coordina la sintesis y montaje de las
fibrillas de celulosa de la pared celular.
recibe y traduce señales ambientales y
hormonales involucradas con el control
del crecimiento y el desarrollo celular.
Núcleo
El núcleo suele ser la estructura más prominente
de la célula. Tiene dos funciones esenciales:
Controla las actividades celulares
determinando que proteínas son
producidas y cuando son producidas.
Almacena la información para controlar
esas actividades (información genética),
que es trasmitida a la generación siguiente
a través de la división celular.
En las células eucarióticas el núcleo está rodeado
por un sistema de doble membrana denominado
membrana nuclear. Esta posee un número grande
de poros de 30-100 nm de diámetro. Las
membranas exterior e interior están unidas
alrededor del poro. Estos poros no son simples
agujeros sino que tienen una estructura compleja.
Cuando una célula se trata con determinados
medios de tinción es posible observar con el
microscopio óptico finos filamentos y gránulos de
cromatina, que se distingue del nucleoplasma que
llena la cavidad nuclear. La cromatina está
constituída por ADN y proteína del tipo de las
histonas. Durante el proceso de división nuclear la
cromatina se condensa progresivamente hasta que
toma la forma de cromosomas. La información
hereditaria de cada organismo está codificada en
el ADN.
Plastidios
Los plastidios o plástidos, junto con la vacuola y
la pared celular, son componentes característicos
de las células vegetales. Cada plastidio se halla
rodeado por una cubierta formada por dos capas
de membrana. Internamente existe además otro
sistema de membranas inmerso en una sustancia
basal más o menos homogénea que se denomina
estroma.
Los plastidios coloreados o cromoplastos maduros
se clasifican comunmente en base a los pigmentos
que contienen. Los cloroplastos contienen
clorofila y carotenoides. La clorofila es
responsable del color verde que poseen muchos
órganos vegetales. En las plantas superiores los
cloroplastos tienen comunmente una forma
discoide de unos 4-6 µm de diámetro. Una sola
célula de mesófilo (la porción interior de una hoja)
contiene usualmente entre 40 a 50 cloroplastos.
Un milímetro cuadrado puede contener así
alrededor de 500.000 cloroplastos. La estructura
interna de los cloroplastos es bastante compleja.
El estroma se encuentra atravezado por un sistema
de membranas aplanadas como sacos que se
denominan tilacoides, que probablemente
consituyen un sólo sistema interconectado. En
ciertas zonas los tilacoides están ordanados en
grana (singular granum), que son como pilas de
monedas que atraviezan el estroma.
Los cloroplastos son orgánulos semiautónomos
que se parecen a bacterias en muchos aspectos.
Por ejemplo poseen ADN formando anillos,
ribosomas de tamaño similar a los de las bacterias.
En cierta medida los cloroplastos son capaces de
sintetizar parte de sus componentes, aunque la
mayoría están codificados en el núcleo, son
sintetizados en el citoplasma y luego importados
al cloroplasto. Los cloroplastos son el sitio donde
se realiza la fotosíntesis y son lugar de
almacenamiento temporario de almidón. Además
de glucosa fruto de la fotosíntesis, en los
cloroplastos se fabrican también aminoácidos y
ácidos grasos.
Los cromoplastos son plastidios pigmentados,
que usualmente han perdido la clorofila pero han
retenido carotenoides. Son los responsables de
colores vistosos en muchas flores, hojas
senescentes y frutos.
Leucoplastos son plastidios no pigmentados.
Algunos sintetizan almidón y son denominados
amiloplastos. Otros sintetizan proteinas o aceites.
Cuando son expuestos a la luz los leucoplastos
pueden transformarse en cloroplastos.
Proplastidios son plastidios pequeños, incoloros o
verde pálido presentes en órganos en crecimiento,
que sirven de precursores a los demás tipos de
plastidios. Los plastidios tienen una cierta
capacidad de transformarse unos en otros

8 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
(interconvertibilidad) según las condiciones del
medio en que está situado el órgano en cuestión.
Mitocondria
Como en el caso de los plastidios, las
mitocondrias se halla envueltas en dos
membranas. La membrana interna se encuentra
plegada formando sacos o invaginaciones que se
denominan crestas, que incrementan
notablemente la superficie interna disponible para
la instalación de enzimas y para las reacciones
asociadas a estas. La mitocondrias son de menores
dimensiones que los plastidios (0.5 µm de
diámetro), y su forma y longitud es bastante
variable. En general las mitocondrias son apenas
visibles al microscopio óptico. Las mitocondrias
son el sitio primordial de la respiración.
Estudios microfotográficos han mostrado que las
mitocondrias se encuentran en un movimiento
bastante activo, fusionandose y dividiendose. Las
mitocondrias tienden a migrar y congregarse en
los lugares de la célula donde es necesaria energía.
Por ejemplo en las células flageladas suelen
agruparse en la base de los flagelos.
Como los plastidios, las mitocondrias son
orgánulos semiautónomos, que contienen
elementos necesarios para la síntesis de algunos de
sus componentes: ARN, ADN y ribosomas
similares a los de las bacterias.
Origen evolutivo de cloroplastos y
mitocondrias
Debido a la similitud entre las bacterias, y los
cloroplastos y las mitocondrias de las células
eucarióticas, parece muy problable que ambos
tipos de orgánulos se originaron como bacterias
que encontraron abrigo en células heterotróficas
grandes. Este proceso debió haberse producido en
etapas tempranas de la evolución. Estas células
bacterianas más pequeñas obtuvieron
probablemente un refugio, un ambiente más
estable en la célula hospedante mayor, que a
cambio recibió entidades capaces de producir
energía más eficientemente. Este proceso de
asociación y adaptación mutua puede designarse
como simbiosis. Esta nuevo tipo de célula pudo
adaptarse mejor que las primitivas formas a otros
ambientes del pasado.
Microcuerpos
Los microcuerpos, a diferencia de plastidios y
mitocondrias, están rodeados por una membrana
simple. Tienen un tamaño de 0.5-1.5 (m. Carecen
de ribosomas y ADN, y deben importar todos sus
componentes. Están asociados frecuentemente al
retículo endoplasmático. Un tipo especial de
microcuerpos, denominados peroxisomas, tienen
un papel muy importante en algunos ciclos
bioquímicos que se llevan a cabo en la célula,
como el ciclo del ácido glicólico. Otro tipo
especial son los glioxisomas, que están asociados
a los cloroplastos y mitocondrias en el ciclo del
glioxilato, necesario para la transformación de
grasas en carbohidratos.
Vacuola
Junto con la presencia de plastidios y pared
celular, la vacuola es uno de los tres elementos
característicos que distinguen a las células
vegetales de las animales. Las vacuolas son
regiones rodeadas por una membrana y llenas una
solución liquida denominada savia celular. La
membrana que rodea a las vacuolas se denomina
tonoplasto, o membrana vacuolar. En una célula
puede haber distintas vacuolas con diferentes
funciones. En especial las células pequeñas tienen
varias vacuolas que usualemente se fusionan en
una sola vacuola central a la madurez. En una
célula madura frecuentemente el 90% del volumen
total de la célula puede estar ocupado por la
vacuola. La mayor parte del incremento de
volumen celular durante el crecimiento se debe a
incremento de volumen vacuolar. Una de las
principales consecuencias de la presencia de una
gran vacuola es el desarrollo de la presión de
turgencia, y del mantenimiento de la rigidez de los
tejidos.
El principal componente del jugo vacuolar es
agua. Los otros componentes varían de acuerdo al
tipo de célula, tipo y estado fisiológico de la
planta. En la vacuola se almacenan sustancias de
reserva (ej. sacarosa), pigmentos (ej. los
pigmentos de las flores llamados antocianinas).
Se piensa que la vacuola se origina a partir del
retículo endoplasmático asociada al retículo de
Golgi, en forma similar a los lisosomas de las
células animales.
Ribosomas
Los ribosomas son partículas de tamaño pequeño,
de unos 17 a 23 nm de diámetro, compuestos de
proteína y ARN en proporciones más o menos
iguales. Los ribosomas son los sitios donde los
aminoácidos de asocian en cadenas para formar

9 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
proteínas. Los ribosmas se presentan con
frecuencia asociados al retículo endoplasmático,
que recibe en ese caso el nombre de reticulo
endoplasmático rugoso. Los ribosomas
involucrados en la síntesis activa de proteínas se
encuentran con frecuencia asociados en cadenas
denominadas poli-ribosomas.
Retículo endoplasmático
El retículo endoplasmático (RE) es un complejo
tridimensional de membranas de extensión
indefinida. Visto en sección, el RE aparece como
dos membranas paralelas con un espacio
transparente y estecho que las separa, llamado
lumen. El RE se divide en RE rugoso, cuando se
encuentra asociado a ribosomas, y RE liso, cuando
carece de ribosomas.
El RE forma un sistema de vesículas
interconectadas que canaliza sustancias hacia
distintas partes de la célula. Filamento del RE
atraviesan a células contiguas a través de los
plasmodesmas. El RE es también el principal
lugar de síntesis de membranas de la célula.
Aparato de Golgi
El término aparato de Golgi se aplica
colectivamente al conjunto de orgánulos
denominados dictiosomas. Los dictiosmas son
complejos de cisternas aplanadas que usualmente
se ramifican en su periferia y originan cuerpos
globulares. Las células de las plantas superiores
tienen dictiosomas compuestos por unas 8 de
estas membranas superpuestas. Los dicitiosomas
presentan dos caras, en una de las cuales tiene
lugar la formación de las cisternas, mientras que
en la opuesta la maduración de las mismas.
Los dictiosomas se encuentran involucrados con
procesos de secreción, en particular de sustancias
constituyentes de la pared celular. Algunos
compuestos celulares son sintetizados por el RE
para luego pasar el aparato de Golgi donde son
modificados previamente a la secreción.
El concepto de endomembranas
Las membranas de la célula son estructuras
dinámicas, móviles, que cambian constantemente
de forma y area superficial. Según el concepto de
interrelación de las endomembranas, todas las
membranas citoplasmáticas (a excepción de las
membranas de mitocondrias y plastidios), forman
un sistema continuo, interconectado. De acuerdo a
esta idea, el RE y el aparato de Golgi forman una
unidad funcinal. Si bien todas las membranas
celulares persisten en el tiempo, sus componentes
moleculares están siendo continuamente
reciclados a traves del sistema RE y Golgi.
Citoesqueleto
Las células eucarióticas poseen un citoesqueleto,
una red compleja de filamentos proteicos que se
extiende internamente por el citoplasma. El
citoesqueleto está estrechamente ligado a varios
procesos, como por ejemplo la división celular, el
crecimiento y elongación celular, el movimiento
de orgánulos en el interior de la célula.
El citoesqueto consiste en dos tipos de filamentos
proteicos: microtúbulos y microfilamentos. Los
microtúbulos son estructuras largas, cilíndricas de
unos 24 nm de diámetro y de longitud variable.
Están formados por subunidades de proteina
denominada tubulina. Los microtúbulos son por
ejemplo responsables de transportar las vesículas
de los dictiosomas hacia la pared, y de ordenar las
microfibrillas de celulosa cuando son depositadas
en la pared celular. Los microfilamentos están
compuestos de una proteína llamada actina y están
frecuentemente asociados a los microtúbulos e
involucrados en procesos similares.
Sustancias ergásticas
Las sustancias ergásticas son los productos
pasivos de la actividad celular. Algunas de estas
sustancias son productos almacenados, mientras
que otras son productos de desecho. Ejemplos de
sustancias ergásticas son : almidón, cristales,
antocianinas, resinas, gomas, taninos , cuerpos
proteicos, gotas lipídicas.
Las sustancias ergásticas se encuentran en la pared
celular, en el citosol, en los orgánulos y en la
vacuola.
Se habla de metabolítos primarios cuando se trata
de sustancias que juegan un papel básico en el
metabolismo y son con frecuencia reutilizados
(almidón). Metabolitos secundarios son aquellos
que no tienen aparentemente un papel esencial en
la actividad celular primaria (ej. alcaloides).
PARED CELULAR
La pared celular es una de las principales
características que diferencian las células

10 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
vegetales de las células animales. Su presencia es
la base de muchas de las características de los
vegetales como organismos. La pared vegetal
limita el tamaño del protoplasma, determina su
forma, y previene la ruptura de la membrana
plasmática cuando el protoplasma se expande
debido a la absorción agua.
En el pasado la pared celular era considerada un
elemento exterior (una "excreción"), en cambio
actualmente se le reconoce funciones esenciales y
específicas. La pared celular contiene una
variedad de enzimas y juega un rol importante en
la absorción, transporte y secreción de sustancias.
También pueden servir como el lugar de actividad
digestiva o lisosómica. Además, un papel activo
en la defensa contra patógenos, bacterias y
hongos. Ella puede recibir y procesar la
información de la superficie del patógeno, y
transmitirla al protoplasma. El protoplasma puede
reaccionar volviendose resistente al ataque ya sea
produciendo fitoalexinas (sustancias antibióticas
tóxicas para los patógenos), o desencadenando la
síntesis de sustancias como lignina y suberina, que
actúan como barreras a la invasión.
Componenetes químicos
El componente más típico de las paredes celulares
vegetales es la celulosa. La celulosa es un
polisacárido estructural cuyo monómero es la
glucosa. Las moléculas de celulosa, que son largas
y delgadas, se unen entre si formando micelas.
Estas a su vez se agrupan en microfibrillas, de
unos 10-25 nanómetros (nm), visibles a
microscopio electrónico. La microfibrillas se
reúnen en macrofibrillas que ya son visibles al
microscopio óptico. Estas tienen alrededor de 0.5
micrómetros (µm) de diámtro y 4 mm de largo.
Las moléculas de celulosa reunídas de este modo
tienen una resistencia a la tracción que supera a la
que tiene una fibra de acero de un diámetro
equivalente. Este ordenamiento en fibras le otorga
elasticidad.
La estructura de celulosa está "embebida" en una
matriz de moléculas no celulósicas. Algunas de
estas moléculas son polisacáridos llamados
hemicelulosas y sustancias pécticas o pectinas que
están quimicamente emparentadas.
Otro componente muy importante en las paredes
celulares en muchos tipos de células es la lignina,
que después de la celulosa es la sustancia más
abundante que se encuentra en los vegetales. A
diferencia de la celulosa, la lignina le otorga
rigidez a la pared, y por lo tanto, se encuentra en
paredes de células que cumplen una función
mecánica.
Cutina, suberina y ceras son sustancias grasas que
se encuentran en las paredes celulares de las
células ubicadas en tejidos exteriores del cuerpo
vegetal. La cutina se encuentra en las paredes
epidérmicas. La suberina se encuentra en las
cortezas corchosas. Ambas sustancias suelen
presentarse conjuntamente con ceras, cuya función
es reducir la pérdida de agua.
En algunos grupos vegetales, además de
sustancias orgánicas, en la pared celular se
depositan sustancias inorgánicas, como sílice o
carbonato de calcio. Depósitos de sílice son
comunes en las epidermis de las gramíneas,
ciperáceas y equisetáceas. Carbonato de calcio se
encuentra en distintos grupos de algas y en pelos
epidérmicos de cucurbitáceas.
Capas de la pared.
Las paredes celulares difieren marcadamente en su
espesor, que varía en relación al rol que cada
célula cumple dentro de la estructura de la planta.
Las capas celulósicas que se forman inicialmente
reciben el nombre de pared primaria. La zona de
unión entre las paredes primarias de dos células
adyacentes se denomina laminilla o lámina
media. Posteriormente pueden depositarse varias
capas más, hacia el interior de la célula, que
forman la pared secundaria.
Laminilla media
La laminilla media está constituída por sustancias
pécticas. Muchas veces es difícil distinguir la
laminilla media de la pared primaria, en particular
en células que han desarrollado paredes
secundaria engrosadas. Cuando una pared celular
se lignifica, este proceso se inicia en la laminilla
media, progresando paulatinamente hacia la pared
primaria y posteriormente la secundaria. El
colorante rojo de rutenio tiñe específicamente la
laminilla media.
Pared primaria
La pared primaria se deposita antes y durante el
crecimiento de la célula. Además de celulosa,
hemicelulosas y pectina, la pared primaria tiene
enzimas y glicoproteínas. Comunmente, las
células que se dividen activamente o aquellas que
tienen gran actividad metabólica (fotosíntesis,

11 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
respiración, excresión) poseen sólo pared
primaria. Estas células, es decir células vivas sin
pared secundaria son capaces de perder su
especialización, dividirse y transformase en otros
tipos de células (dediferenciación). Estas células
son las pueden involucrarse activamente en la
regeneración de nuevos órganos o en la
cicatrización de heridas.
Usualmente la pared primaria no tiene un grosor
uniforme, sino que posee áreas más delgadas
denominadas campos de punteaduras primarias.
Estas zonas son normalmente atravesados por
cordones de citoplasma que se denominan
plamodesmas.
Pared secundaria
Muchas células vegetales sólo poseen pared
primaria. En cambio en otras se deposita en el
interior de la pared primaria una capa de pared
secundaria. La pared secundaria se forma
normalmente en células que han detenido su
crecimiento. La paredes secundarias tienen gran
importancia en las células que tienen función de
sostén y de conducción de agua. En estas células,
el protoplasto suele morir luego que se ha
depositado la pared secundaria. La celulosa suele
ser más abundante en las paredes secundarias que
en la primarias y no hay pectinas. Tampoco se
encuentran enzimas ni glicoproteínas. Con
frecuencia se distinguen tres capas en la pared
secundaria, exterior, media e interna. Estas capas
difiren en la orientación de las microfibrillas de
celulosa. Esta estructura laminar incrementa en
modo significativo la resistencia. Lignina suele
agregarse a la estructura celulósica en las células
de los troncos de árboles.
La pared secundaria no se deposita en las zonas
con punteduras, lo que da como resultado
depresiones muy típicas que se observan al
microscopio.
Crecimiento de la Pared Celular
La pared celular crece tanto en espesor como en
superfice. El crecimiento es un proceso complejo
controlado por el protoplasma, que requiere el
ablandamiento de las estructuras presentes
conjuntamente con un incremento de la síntesis de
proteína y de la respiración que aporta energía en
el proceso, y aumento de la absorción de agua. El
crecimiento de la pared esta influenciado por
hormonas vegetales, en especial las llamadas
auxinas. La mayor parte de las nuevas
microfibrillas se ubican encimas de las formadas
previamente (aposición), pero algunas pueden
insertarse dentro de la estructura preformada
(intususcepción).
En células que crecen en todo sentido las fibrillas
se disponen al azar, formando una red irregular.
Este tipo de células se encuentra en la médula de
los tallo, en tejidos de almacenamiento o en
tejidos cultivados in vitro. En cambio, en células
que se alargan en un sentido principal las fibrillas
se ordenan transversalmente al eje de
alargamiento, y a medida que el alargamiento
tiene lugar, pueden quedar en una posición casi
paralela a este eje. La síntesis de las microfibrillas
de celulosa no está del todo aclarada pero se sabe
que son sintetizadas por enzimas inmersas en la
membrana plasmática y su orientación es
controlada por microtúbulos presentes en el
interior de esta membrana. Las hemicelulosas y
pectinas que forman la matriz son depositadas por
dictiosomas. En células en crecimiento la síntesis
de pectinas es predominante, mientras que en
células que no crecen el balance favorece a las
hemicelulosas.
Plasmodesmas
Los protoplastos de células adyacentes están
conectados por plasmodesmas. Si bien las
perforaciones en la pared celular son visibles con
el microscopio óptico, su estructura detallada sólo
se pudo conocer bien con el empleo del
microscopio electrónico. Con este instrumento se
puede ver que los plasmodesmas son canales
delgados (30-60 nm de diámetro), rodeados de
membrana plasmática y atravezados por un tubo
de retículo endoplasmático denominado
desmotúbulo. Muchos plasmodesmas se forman
durante la división celular.
Espacios intercelulares
Las células suelen tener toda o una proporción
sustancial de su pared en contacto con otras
células. Sin embargo en ciertos tipos de tejidos
existen espacios intercelulares o meatos. Estos
espacios son en algunos casos muy importantes
para el funcionamiento celular. Por ejemplo las
células fotosintéticas tienen una proporción grande
de su superficie en contacto con espacios
intercelulares por los cuales se difunden O2 y CO2.

12 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
3.1. BRIOFITAS Algas verdes, briofitas y plantas vasculares
Las plantas terrestres se originaron a partir de las
algas verdes (Chlorophyta). Existen características
citológicas y bioquímicas que sustentan este
parentezco, compartido por briofitas, plantas
vasculares y algas verdes:
almidón como principal sustancia de reserva
almidón formado y depositado en los
plastidios
celulosa como principal componente de la
pared
formación de un fragmoplasto y placa celular
durante la mitosis.
Las briofitas fueron las primeras plantas
verdes en colonizar la tierra, hace unos 420
millones de años. Tres grupos de plantas:
hepáticas (división Hepatophyta), antocerotes
(división Anthocerophyta) y musgos (división
Bryophyta) han sido denominados "briofitas" en
sentido amplio. Sin embargo el parentezco preciso
entre los tres no está claro y forman un conjunto
parafilético (fig. 3.1.11). Durante su ciclo de vida
(fig. 3.1.10) presentan alternancia de
generaciones, y su característica común es que el
gametofito es la generación dominante,
nutricionalmente independiente del esporofito,
mientras que este último está adosado al primero y
depende toda su vida de él. Por el contrario, en
todos los grupos de plantas vasculares el
esporofito es la fase conspicua y dominante. En el
gametofito se producen las gametas femeninas en
gametangios llamados arquegonios, y las
masculinas en anteridios. Ambos gametangios
están cubiertos por capas de células estériles que
protegen a las gametas.Algunas especies
desarrollan estructuras especializadas portadoras
de arquegonios o de anteridios (arquegonióforos y
anteridióforos, (fig. 3.1.1). En el esporofito se
producen esporas haploides en esporangios o
cápsulas.Las hepáticas son consideradas como las
briofitas más sencillas, pues carecen de células
conductoras especializadas (aunque se han
encontrado en unos pocos grupos) y de estomas.
Los antocerotes carecen de células conductoras,
sin embargo poseen estomas, una característica
común con todas las plantas vasculares. Los
musgos a su vez poseen células conductoras de
agua y de nutrientes en sus gametofitos y
esporofitos (aunque no todos los géneros los
poseen).
Hepáticas
Las hepáticas son un grupo de unas 8000 especies
de plantas pequeñas y usualmente inconspicuas,
aunque pueden formar masas muy densas en
ambientes favorables, comunmente húmedos y
sombríos. El nombre hepática fue dado en la edad
media porque el talo de algunas especies tiene un
parecido con los lóbulos hepáticos.
Hepáticas talosas: se denomina así a un grupo
que se caracteriza por tener gametofitos
aplanados, con simetría dorsiventral, ramificados
dicotómicamente, con una línea media y rizoides
que la fijan al sustrato. Las hepáticas pueden
reproducirse vegetativamente por fragmentación.
También suelen producir estructuras
especializadas en forma de copa (conceptáculos)
donde se producen cuerpos multicelulares
llamados yemas, (fig. 3.1.2). Estas son dispersadas
principalmente por gotas de lluvia.
Figura 3.1.1.Marchantia sp. con anteridióforos (A) y
arquegonióforos inmaduros (B).
Figura 3.1.2. Talo de Marchantia sp. con conceptáculos,
estructuras de reproducción vegetativa o yemas.
Hepáticas foliosas: grupo más numeroso que las
talosas, llamadas así debido a que su talo adquiere

13 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
aspecto de hojitas. Las hojas (o “filoides”) están
constituídas por una capa uniestratificada de células
que se disponen normalmente en tres hileras sobre
un eje central: dos dorsales que se insertan
diagonalmente y una ventral de insersión
transversal.
Figura 3.1.3. Plagiochila asplenioides, hepática foliosa.
Antocerotes
Los gametofitos de los antocerotes se asemejan al
de las hepáticas talosas. Sin embargo hay varios
caracteres que indican un parentezco bastante
lejano entre ambos grupos. Por ejemplo cada
célula de un antocerote tiene un único cloroplasto
(como muchas algas verdes) en lugar de varios
cloroplastos discoides, como en las hepáticas y
plantas superiores. Cada cloroplasto tiene un
pirenoide, lo cual hace la similitud con ciertas
algas verdes mayor aún. Por otro lado la presencia
de estomas hace pensar en cierta afinidad con las
plantas vasculares.
Figura 3.1.4. Gametofito de antocerotes con varios
esporofitos de color verde más claro emergiendo.
El gametofito de los antocerotes es
marcadamente dorsiventral, con aspecto de roseta,
de 1-2 cm de diámetro. En el gametofito de
muchos antoceros, aparecen notables cavidades ,
que se encuentran llenas de mucílago y
frecuentmente habitadas por algas cianofíceas del
género Nostoc, que fijan nitrógeno. El esporofito
de los antoceros tiene un pie inmerso en el
gametofito, y un esporangio o cápsula emergiendo
de él, con una región meristemática intermedia
que genera un crecimiento activo siempre que las
condiciones sean favorables. El esporangio se abre
por el extremo en dos valvas, liberando
paulatinamente las esporas.
Musgos
Los musgos constituyen un grupo diverso de unas
10.000 especies. Son usualmente abundantes en
ambientes húmedos. En ciertos casos los musgos
pueden dominar totalmente grandes extensiones
de terreno, particularmente en regiones
montañosas y circumpolares. Sin embargo algunas
especies son capaces de vivir en ambientes áridos.
Varias de éstas tienen la capacidad de
deshidratarse completamente, permanecer en
estado latente por años y reactivarse en pocos
minutos una vez rehidratados.
El gametofito de los musgos es
inicialmente filamentoso. Este filamento se
denomina protonema, que se genera a partir de
una espora que germina. El protonema produce
estructuras como yemas, a partir de las cuales
crece un gametofito folioso. Los gametofitos
pueden variar en complejidad y tamaño, 0.5 mm a
50 cm de longitud.
El gametofito es usualmente erecto y con
simetría radial, no aplanado y dorsiventral como
las hepáticas talosas. Posee estructuras análogas a
hojas y tallos y rizoides que solo lo fijan al
sustrato. Las hojas son usualmente formadas por
una sola capa de células. Los tallos poseen un haz
central de células denominadas hidroides. Estas
son células alargadas, carentes de protoplasma,
especializadas en el tranporte de agua y solutos,
que carecen de engrosamientos en la pared celular
como las tráqueas y traqueidas de las plantas
vasculares. En algunos grupos existen células
especializadas en la conducción de sustancias
elaboradas, denominadas leptoides.
Figura 3.1.5. “Césped” de Polytrichum sp. con los
gametofitos de color verde intenso y. los esporofitos
con seda y cápsula de color rojo y naranja

14 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Figura 3.1.6. Polytrichum sp. Gametofitos de color verde
intenso y esporofitos con cápsula.
Figura 3.1.7. Polytrichum sp. Cápsula cubierta por la
caliptra (izquierda), cápsula abierta sin opérculo (centro),
cápsula que mantiene todavía su opérculo (derecha).
En el extremo del eje principal o de ramas
laterales se producen anteridios y arquegonios.
Los esporofitos poseen una base o pie inmersa en
el gametofito y una estructura alargada
denominada seta, en el extremo de la cual se halla
una cápsula, donde se producen las esporas por
meiosis. Las cápsulas de los musgos se abre
comunmente por cambios en la humedad
atmosférica. La forma de apertura de la cápsula es
variada y uno de los criterios empleados en la
clasificación de los distintos grupos de musgos.
Relevancia ecológica y económica
de las briofitas
En las regiones circumpolares los musgos, en
particular del género Sphagnum (Figura 3.1.9),
son capaces de dominar totalmente el paisaje. Se
estima que estos ambientes cubren alrededor del
1% de la superficie terrestre, una extensión
superior a toda superficie de Argentina. Buena
parte de esta extensión son turberas, áreas
pantanosas, ácidas, cubiertas por una capa que puede
llegar a tener varios metros de espesor de musgos.
La capa superior es una cubierta delgada de musgos
vivos, sobre restos de materia muerta. La turba, una
vez seca puede ser empleada como combustible.
También se usa extensamente en jardinería, para la
preparación de suelo artificial y relleno de macetas,
pues tiene la capacidad de retener 20 veces su peso
en agua, manteniendo a la vez una buena aireación.
Las "barbas del monte" del NOA, los musgos que
cubren los árboles en el bosque montano inferior,
pueden ser usados para el mismo fin.
En los ambientes de bosques y selvas, las
briofitas y en particular musgos y hepáticas
foliosas, cumplen un rol muy importante
formando un sustrato sobre las superficies de
rocas y troncos, donde se pueden establecer otras
plantas. Por otro lado esta cubierta de briofitas
cumple un papel importante interceptando
“precipitaciones horizontales”, es decir captando
niebla y humedad aumentando el total de agua que
llega al suelo. Este fenómeno es muy importante
en las selvas nubladas del mundo como por
ejemplo las Yungas.
Figura 3.1.8. Tronco cubierto de briofitas en la sierra de
San Javier,en el piso de selva montana de Yungas.
Las briofitas son especialmente sensibles a
la contaminación ambiental y atmosférica, y
suelen estar ausentes en las grandes ciudades. Por
su sensibilidad pueden ser empleadas como
indicadores biológicos de contaminación.

15 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Ciclo biológico de una briofita
gametofitos adosados al sustrato por medio de
células o filamentos de células alargados
denominados rizoides; estos rizoides sirven
principalmente para fijar al sustrato, ya que la
absorción de agua y sustancias inorgánicas se
produce directamente a través de toda la
superficie del gametofito.
la célula sexual masculina (anterozoide o
espermatozoide) es flagelado y debe nadar en
agua para llegar hasta la célula sexual femenina
y fertilizarla.
la célula sexual femenina (ovocélula) se
encuentra en una estructura con forma de
botella, denominada arquegonio, con un cuello
alargado y una porción basal engrosada, el
vientre. Las células del canal del cuello se
desintegran cuando el arquegonio está maduro,
dando como resultado un canal lleno de fluido a
través del cual nadan los anterozoides.
Los anterozoides se originan en estructuras con
forma de clava, denominadas anteridios,
constituídas por una capa unicelular que encierra
numerosas células espermatógenas.
Luego de la fertilización la cigota se desarrolla
formando un embrión. Cuando éste se desarrolla
formando el esporofito, las células del arquegonio
crecen conjuntamente encerrándolo por un tiempo,
para luego romperse; los restos del arquegonio
agrandado durante este proceso recibe el nombre de
caliptra.
Figura 3.1.10. Ciclo biológico de un musgo.

16 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Figura 3.1.11. Esquema de las probables relaciones filogenéticas entre los distintos grupos de briofitas y las
restantes plantas terrestres, y su denominación actual (parte superior). Sobre la rama inferior aparecen los
distintos pasos evolutivos y la terminología clásica empleada para agrupar a las briofitas en sentido amplio y
a las plantas vasculares sin semillas.
Figura 3.1.12. Dendroligotrichum dendroides, el
“musgo pinito”, uno de los musgos de mayor
tamaño presente en la selva valdiviana de Chile y la
isla sur de Nueva Zelanda, regiones de lluvias muy
elevadas.
Figura 3.1.13. Mnium sp., un género de musgos que
se destaca por sus hojas notablemente desarrolladas,
formadas por una sola capa células.

17 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
3.2 PLANTAS VASCULARES SIN
SEMILLALas plantas vasculares o traqueofitas presentan
tejidos de conducción altamente especializados,
con paredes reforzadas con lignina, lo que mejora
la capacidad de conducción de agua, y a la vez les
otorga rigidez estructural, permitiendo la
formación de órganos altamente complejos y de
gran tamaño. Además del sistema vascular las
traqueofitas presentan tejidos epidérmicos de
aislamiento y un sistema de raíces absorbentes,
elementos estos que combinados han permitido la
colonización extensiva de los ambientes terrestres.
A diferencia de las briofitas, en las traqueofitas la
fase dominante es la fase diploide o esporofítica.
En la actualidad las traqueofitas se separan
filogenéticamente en dos grandes grupos: las
licofitas y las eufilofitas (“plantas con verdaderas
hojas”), nombre que hace referencia a que sus
hojas generalmente son desarrolladas y con una
estructura compleja de nervios conductores. A su
vez las eufilofitas se subdividen en monilofitas
(los helechos, incluyendo también Equisetaceae y
Psilotaceae) y espermatofitas, (“plantas con
semillas”). En este texto usamos el término
helecho como sinónimo de monilofita. En el
pasado se usó (y algunos aún lo hacen) el término
“helecho” o “pteridofita” en un sentido más
amplio, para denominar a todas las plantas
vasculares sin semillas, es decir el conjunto de
licofitas y monilofitas. Agrupados de esta manera,
los “helechos” o “pteridofitas” constituyen un
grupo parafilético.
Las hojas responden a dos tipos generales:
micrófilos, órganos de estructura sencilla con una
sola nervadura y usualmente de tamaño reducido,
y macrófilos (=megáfilos), estructuras complejas,
con un sistema de nervaduras bien desarrollado,
que les permite en muchos casos alcanzar grandes
dimensiones.
Las licofitas y monilofitas pueden presentar dos
tipos de esporangios: eusporangios, recubiertos
por dos o más capas de células a la madurez, o
más comúnmente leptosporangios, recubiertos
con una sola capa de células. En los esporangios
están presentes células madres, que por meiosis
generan esporas. Estas esporas son dispersadas
usualmente por el viento, y cuando germinan
generan un protalo haploide, que puede ser
fotosintético o simbiótico con hongos del suelo.
Tanto licofitas como monilofitas presentan
especies que producen un solo tipo de esporas,
denominadas isosporadas (u homosporadas), o
especies que producen dos tipos de esporas, es
decir heterosporadas. Estas dos clases de
organismos presentan en consecuencia ciclos
biológicos claramente diferentes. Tanto licofitas
como monilofitas producen anterozoides
flagelados. Sin embargo, mientras los de las
licofitas son biflagelados, lo que indicaría un
parentezco más cercano a las briofitas, las
monilofitas los poseen multiflagelados.
Ciclos biológicos
Ciclo isosporado (fig.3.2.12): El esporofito posee
esporangios, sobre sus hojas o en estructuras
especiales. Las esporas son liberadas por el
esporangio y germinan generando un protalo
haploide, en la mayoría de los casos con un
aspecto similar a una hepática talosa. En el protalo
se forman anteridios y arquegonios. Los
anteridios liberan anterozoides que en presencia
de gotitas de agua nadan hasta los arquegonios,
orientados quimitrópicamente, y fertilizan a la
ovocélula (u oosfera), produciendo un zigoto
diploide. El zigoto se desarrolla como un
esporofito, que posee esporangios donde por
meiosis se producirán las esporas haploides para
reiniciar el ciclo.
Ciclo heterosporado (fig. 3.2.13): poseen
macrosporangios y microsporangios. Los
primeros producen microsporas que en su interior
producen un gametofito masculino que produce un
anteridio. Este libera anterozoides flagelados que
nadan hacia el gametofito femenino. El
macrosporangio produce macrosporas, en cuyo
interior se genera un gametofito femenino con
arquegonios. Dentro del arquegonio se realiza la
fecundación de la ovocélula por parte de un
anterozoide, generándose un zigoto diploide. El
zigoto se desarrolla en un esporofito, que a la
madurez produce macro y microsporangios.
Licofitas
Las licofitas emergieron hace unos 400 millones
de años, como los primeras plantas vasculares en
la superficie terrestre. Su máxima expansión fue

18 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
en el período Carbonífero (hace 345-290 millones
de años), cuando como organismos de gran
tamaño, con tallos de hasta 40 m de alto y 2 m de
diámetro, dominaron la tierra. Alrededor de la
mitad de los yacimientos de carbón de piedra del
mundo se originaron a partir de estas licofitas
ancestrales.
Figura 3.2.1.Lycopodium clavatum
(Lycopodiaceae), licofita con estróbilos terminales
de las Yungas de Jujuy y Salta.
Figura 3.2.2. Huperzia saururus (Lycopodiaceae),
licofita de los pastizales de neblina de NOA, entre
los 2000 y 3700 m. No forma estróbilos y los
esporangios pueden aparecer en la axila de
cualquier hoja normal.
Las licofitas modernas son en cambio
relativamente pequeñas, de porte herbáceo, que
poseen micrófilos, con esporangios ubicados en
las axilas de éstos. Existen actualmente algo más
de 1000 especies, separadas en tres familias:
Lycopodiaceae, Sellaginelaceae e Isoetaceae.
Las Licopodiaceae son isosporadas, mientras que
las Selaginellaceae e Iisoetaceae son formas
heterosporadas.
Figura 3.2.3. Sellaginela nova-hollandiae
(Sellaginelaceae), común en bosques de aliso.
(Abajo) Detalle de Eje caulinar con estróbilo
terminal (E) y micrófilo (M). Tomado de Boero
(1993).
Figura 3.2.4. Isoëtes alcalophila (Isoetaceae), es
endémica de las lagunas de Huaca Huasi (4300 m),
Cumbres Calchaquíes, provincia de Tucumán,
donde forma céspedes sumergidos.

19 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Monilofitas o helechos
Los helechos dominaron los ambientes terrestres
luego de las licofitas, hace unos 300 millones de
años, pero fueron paulatinamente desplazadas por
las espermatofitas. La gran mayoría de las formas
actuales están adaptadas a coexistir en ambientes
dominados por aquellas. Presentan una diversidad
morfológica mucho mayor que las licofitas. La
mayoría tienen tallos subterráneos, frecuentemente
rizomas, con un penacho de hojas, usualmente
compuestas o profundamente lobadas, con
numerosas nervaduras y que pueden alcanzar
grandes dimensiones (hasta varios metros en los
helechos arborescentes). Las hojas reciben el
nombre de frondes. Los folíolos o lóbulos de las
frondes son denominados pinas. También existen
helechos con hojas simples o relativamente
sencillas. Dos familias, las Psilotaceae y
Equisetaceae, las tienen muy reducidas, con una
sola o ninguna nervadura, es decir micrófilos, más
parecidas a las de las licofitas. Sin embargo
actualmente se acepta que estos dos grupos son
helechos, altamente modificados
morfológicamente. Los helechos incluyen unas 40
familias, distribuidas en alrededor de 300 géneros
y 9000 especies.
La mayoría de los helechos son isosporados,
produciendo un solo tipo de esporas. Al germinar
estas producen un gametofito, generalmente
bisexual, que produce los dos tipos de gametas,
oosferas en arquegonios y anterozoides en
anteridios. Por la unión de anterozoide y oosfera
se genera un zigoto. La presencia de esporas
permite a los helechos ser muy eficientes en la
dispersión a larga distancia por viento. En la
mayoría de los helechos los esporangios se
encuentran agrupados en conjuntos llamados
soros, que pueden o no estar recubiertos por una
membrana protectora de origen foliar, el indusio.
La forma y posición de los soros son carateres
muy usados para la identificación de los helechos.
Equisetaceae
Los equisetos o “cola de caballo” son plantas con
tallos fotosintéticos, con nudos y entrenudos muy
marcados y hojas muy reducidas. Estas se
encuentran dispuestas en forma verticilada y las
ramificaciones se disponen de la misma manera.
Los esporangios se ubican, en un número de 6-8,
en esporangióforos peltados, que se agrupan a su
vez en estructuras terminales denominadas
“estróbilos”. Los equisetos son buenos
colonizadores de áreas pantanosas, orillas de ríos
y lagunas.
Figura 3.2.5. Equisetum giganteum (Equisetaceae)
Arriba: hojas escamiformes y ramas dispuestas en
verticilos sobre un tallo con nudos y entrenudos
marcados. Abajo: esporangios ubicados en
esporangióforos en estróbilos terminales. Es común
en la llanura tucumana, en banquinas inundadas y
cursos de agua.

20 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Helechos leptosporangiados
Los helechos leptosporangiados constituyen el
grupo más grande y común de helechos.
Marsileaceae y Salviniaceae, agrupadas en orden
Salvineales, a diferencia de los restantes grupos de
helechos, son familias heterosporadas,
produciendo macro y microsporas. Este grupo se
caracteriza además por presentar formas acuáticas
o de ambientes periódicamente inundados.
Figura 3.2.6. Típica prefoliación circinada presente
en los helechos leptosporangiados.
Figura 3.2.7. Adiantum lorentzii (Pteridaceae) es
característico de quebradas húmedas y sombreadas
del NOA. Presenta una nerviación dicotómica bien
desarrollada, y soros reniformes ubicados en el
borde de la cara inferior de las pinas.
Figura 3.2.8. Anemia phyllitidis (Schizeaceae)
especie frecuente en la sierra de San Javier. Las dos
pinas basales de la fronde son erectas y fértiles con
numerosos esporangios, mientras que las pinas
distales son exclusivamente fotosintéticas.
Figura 3.2.9. Marsilea sp. (Marsileaceae) helecho
heterosporado de ambientes acuáticos. Se
caracteriza por poseer frondes con 4 pinas florantes.

21 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Figura 3.2.10. Alsophila odonelliana (Cyatheaceae)
es el único helecho arborescente del NOA. Crece en
las yungas de Jujuy y Salta. Sus estípites pueden
superar los 10 m de altura y sus frondes más de 2 m
de largo.
Licofitas y monilofitas cumplen roles ecológicos
destacados en ambientes húmedos,
particularmente selvas nubladas y bosques
templados, en los cuales a veces dominan el
sotobosque. Desde un punto de vista económico
en cambio su importancia es menor. Algunas
especies se emplean en medicina tradicional. Un
número creciente se usan como ornamentales,
particularmente adecuados como plantas de
interior.
Figura 3.11. Microgramma squamulosa
(Polypodiaceae) epífita muy común sobre árboles
en la ciudad de Tucumán y la sierra de San Javier.
Posee tallos trepadores con numerosas raíces
adventicias que se adhieren fuertemente al tronco
que lo soporta. Las hojas maduras producen en el
envés, numerosos soros no recubiertos por un
indusio, que se ven en tres estadios de desarrollo en
la imagen inferior

22 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Ciclo biológico de un helecho leptosporangiado isosporado
Figura 3.2.12. Ciclo biológico de un helecho leptosporangiado isosporado (Polypodium sp.).
Ciclo biológico de una licofita heterosporada (Selaginella sp.).
Figura 3.2.13. Ciclo biológico de una licofita heterosporada (Selaginella sp.).

23 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
3.3.GIMNOSPERMAS La innovación más importante en la evolución
de las plantas vasculares fue la aparición de las
semillas, estructura que les permitió
convertirse en las dominantes en la mayoría de
los paisajes del mundo, desde hace millones de
años.
Las plantas con semillas, o espermatofitas se
separan en dos grandes grupos: las plantas con
semillas desnudas o gimnospermas, y las con
semillas encerradas en un recipiente o “angio”,
las angiospermas. De éstas, las gimnospermas
fueron las primeras en aparecer en el registro
fósil, ya en el periodo carbonífero, cuando el
paisaje era dominado por la Licofitas
primitivas, hace unos 360 millones de años. Ya
a fines de esa era geológica aparecieron
gimnospermas pertenecientes al grupo que
conocemos como coníferas, con características
esencialmente similares a las actuales.
Todas las gimnospermas se
caracterizan por ser leñosas, en general árboles
o arbustos, raramente trepadoras o apoyantes.
No existen formas herbáceas. Se encuentran
presentes entre los 72º N y los 55º S. Algunas
especies son muy importantes y dominan
grandes extensiones en los bosques templados
y fríos del hemisferio norte. Sin embargo el
número total de gimnospermas vivientes en la
actualidad es comparativamente pequeño, unas
820, distribuidas en alrededor de 15 géneros,
en 4 órdenes: Coniferales, Cycadales,
Ginkgoales y Gnetales.
Las gimnospermas son
heterosporadas. Los microesporofilos (hojas
que llevan los microsporangios) se hallan
usualmente formado estructuras compactas
denominadas estróbilos o conos. Las
microesporas (=polen) se dispersan por el
viento como sucede también, en la mayoría de
los helechos. Los macrosporofilos (hojas que
llevan las macrosporas) con frecuencia
también se encuentran en estróbilos, a veces
notablemente desarrollados, pero también
pueden presentarse en forma solitaria. El
proceso de dispersión del polen desde los
microsporofilos hasta los macrosporofilos se
denomina polinización. La fertilización tiene
lugar de distintas maneras y suele producirse
hasta un año después de la polinización. La
posterior formación de la semilla también lleva
meses o hasta un par de años. Las
gimnospermas pueden ser monoicas o dioicas,
es decir poseer macro y microsporofilos en el
mismo pie o en pies diferentes.
Coníferas
Las coníferas (=Coniferales) constituyen el
grupo de gimnospermas más importantes, tanto
desde el punto de vista ecológico, ya que
dominan grandes extensiones de bosques
templados y fríos, como desde el punto de
vista económico, constituyendo la principal
fuente de madera y pulpa de papel en el
mundo.
Figura 3.3.1. Pinus patula (Pinaceae).
Estróbilos microsporangiados y hojas. El “pino
patula”, o “pino llorón”, originario de las
montañas de México, es frecuentemente
cultivado como especie forestal en el NOA. Se
caracteriza por poseer largas hojas aciculares
péndulas.
Figura 3.3.2. Pinus elliottii (Pinaceae).
Estróbilos ovulados y semillas. Proveniente del
NE de Estados Unidos, es el forestal más
cultivado en el NOA.

24 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Figura 3.3.4. Sequoiadendron giganteum
(Cupressaceae), de la Sierra Nevada,
California. Son los organismos más macizos
del mundo, con 1200 m3, 2000 toneladas de
peso y más de 3000 años. El presente en la foto
es el tercero más grande conocido, de 78 m de
altura y 28 m de circunferencia en la base.
Figura 3.3.3.Cedrus deodara (Pinaceae). El
cedro azul, originario del Himalaya es cultivado
con frecuencia. Como muchas Pináceas posee
hojas aciculares que pueden estar ubicadas sobre
tallos cortos, denominados braquiblastos.
La familia Cupressaceae también tiene
representantes gigantescos en Sudamérica. Se
trata del alerce (Fitzroya cupressoides) de los
bosques andino-patagónicos. El individuo más
alto medido de esta especie tiene 57 m de
altura, en el Parque Nacional Los Alerces. Los
alerces son además los árboles más longevos
de Sudamérica. El más viejo fechado hasta
ahora arrojó una edad de 3662 años.
Figura 3.3.5. Cupressus sempervirens
(Cupressaceae). Estróbilos ovulados y hojas
escamiformes imbricadas. Esta es Cupresácea
más común cultivada en Tucumán, conocida
como ciprés común, originario de Europa.

25 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Figura 3.3.6. Podocarpus parlatorei
(Podocarpaceae). Escama ovulífera (izquierda)
y estróbilos microesporangiados (derecha). El
“pino del cerro” es la única especie de conífera
nativa del NOA. Los óvulos se encuentran
aislados sobre una estructura globosa,
comestible a la madurez, que facilita la
dispersión por aves.
Ginkgoales
Este grupo posee en la actualidad una sola
especie sobreviviente, originaria de la China.
Como muchas otras gimnospermas, posee las
hojas sobre braquiblastos, y hojas ovulíferas
individuales. A diferencia de todas las demás
espermatofitas, sus hojas tienen nerviación
dicotómica y son flabeladas, caracteres sólo
presentes en algunos helechos. También como
los helechos, produce espermatozoides
flagelados que nadan hasta la ovocélula para
fecundarla.
Figura 3.3.6. Ginkgo biloba (Ginkgoaceae).
Braquiblasto con óvulos y hojas.
Cycadales
Como Ginkgo, las Cycadales poseen
espermatozoides flagelados. Sin embargo su
estructura vegetativa es muy diferente,
usualmente con un estípite no ramificado y un
solo eje vegetativo apical. Se parecen a las
palmeras, y también a los helechos
arborescentes. Son plantas de crecimiento
extremadamente lento y varias de ellas están
en riesgo de extinción.
Figura 3.3.7. Cycas revoluta (Cycadaceae),
originaria de Nueva Guinea, es común en
muchos jardines del NOA. Planta entera (arriba),
hoja ovulifera (centro), con aspecto de hoja
normal rudimentaria, con óvulos de color rojo en
la zona basal. Las hojas microesporangiadas
formado estróbilos (abajo) y hojas ovulíferas se

26 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
encuentran en plantas diferentes, es decir son
dioicas.
Gnetales
Algunos botánicos consideran a las Gnetales
cercanas a las angiospermas, pues poseen
microsporofilos y macrosporofilos rodeados de
brácteas, en forma análoga a un perianto.
Además poseen xilema con tráqueas, mientras
que todas las demás gimnospermas poseen
solamente traqueidas. Sin embargo estudios
modernos de ADN y cladística, apuntan a un
parentesco cercano a las pináceas.
Figura 3.3.8. Ephedra sp. Típico arbusto de
Ephedra en un ambiente semiárido de Tilcara,
Jujuy (arriba). Semillas maduras de Ephedra
(abajo). El tegumento seminal es carnoso y
coloreado, adaptado a la dispersión por aves.
El género Ephedra está presente en ambientes
áridos y de montaña de Argentina con
alrededor de 10 especies. Tiene hojas muy
pequeñas y rápidamente caedizas. La
fotosíntesis es realizada exclusivamente por los
tallos, que tienen nudos bien marcados.
Ciclo biológico de una gimnosperma (Pinus
sp.)
(1) El esporofito maduro es árbol o arbusto
leñoso de gran tamaño, que produce (2)
estróbilos o conos microsporangiados
(“masculinos”), que poseen microsporofilos
(escamas poliníferas) portadores de los sacos
polínicos donde se produce la meiosis y se
generan los granos de polen. En el mismo pie
se producen (3) estróbilos ovulíferos, con
macro o megaesporofílos (escamas ovulíferas).
En la escama ovulífera se desarrolla un
macrosporangio que produce una célula madre
de la macrospora (4). Por meiosis se originan
cuatro megasporas de las cuales tres degeneran
y solamente una persiste. A partir de la
germinación de esta megaspora se origina un
protalo multicelular o gametofito femenino o
macrogametofito (5). Este gametofito produce
uno o más arquegonios reducidos que generan
una ovocélula cada uno. Cuando un grano de
polen llega a la micrópila forma un tubo
polínico que crece hasta llegar a la ovocélula
(6) y se produce la fertilización (7). La
fertilización da lugar a la formación de un
zigoto que se desarrolla como un embrión
diploide (2n) asociado a reservas alimenticias
originadas a partir del prótalo en el
macrogametofito (8). Al germinar el embrión,
que posee numerosas hojas cotiledonares, da
lugar a una plántula con numerosas hojas. Esta
planta al crecer origina un nuevo esporofito
que al madurar, puede reiniciar el ciclo.

27 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Ciclo biológico de una gimnosperma (Pinus sp.)
Ciclo biológico de una Gimnosperma

28 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
3.4. ANGIOSPERMAS
Plantas con flores
Las Angiospermas o Plantas con Flores, son
las plantas dominantes en los ambientes
terrestres. Las Angiospermas tienen un
prolongado registro fósil, desde el Cretácico
temprano, unos 135 millones de años atrás.
Es un grupo que ha estado separado de los
restantes grupos de Espermatofitas, las
Gimnospermas, desde hace 280 millones
atrás.
La mayoría de las Angiospermas entran en
dos grandes grupos, las Monocotiledóneas
(plantas con un solo cotiledón y granos de
polen usualmente monosulcados),
constituyen alrededor de un 20%, y las
Dicotiledóneas (plantas con dos
cotiledones y granos de polen usualmente
tricolpados o varientes de ello), que
consitituyen alrededor de un 76%.
Modernamente se acepta que existe
aproximadamente un 4% de las
angiospermas que no pertenecen a ninguno
de estos dos grupos mencionados
previamente. En cambio la mayoría de esas
especies pueden agruparse en un tercer
grupo denominado Magnoliidas. Las
Magnoliidas y otras familias menores, que
tampoco se incluyen dentro de Mono- o
Dicotiledóneas, conservan el mayor número
de caracteres ancestrales o primitivos
dentro de las angiospermas.
Se estima que existen unas 250.000
especies de Angiospermas, distribuidas en
unas 350 familias. Tres de ellas son
notablemente grandes e importantes desde
el punto de vista ecológico y económico:
Asteráceas (=Compuestas) con 23.000
PRINCIPALES DIFERENCIAS ENTRE DICOTILEDÓNEAS Y MONOCOTILEDÓNEAS
Dicotiledóneas Monocotiledóneas
Predominan las plantas leñosas: árboles y
arbustos, pero hay muchas hierbas también. La mayoría son hierbas.
Los tejidos conductores se disponen
formando anillos concéntricos en el tallo. Los tejidos conductores se disponen
dispersos regularmente en tallo.
La raíz suele tener un eje central que se
ramifica (raíz pivotante). Las raíces son fasciculadas.
Las hojas suelen tener pecíolo y sus
nervaduras se ramifican y anastomosan
formando una red.
Las hojas no suelen tener pecíolo y
envuelven al tallo. Sus nervaduras
suelen ser paralelas.
Piezas florales en número de cuatro o cinco,
o múltiplos de cuatro o cinco. Piezas florales en número de tres, o
múltiplos de tres en cada ciclo.
Tabla 3.4.1. Principales diferencias entre Dicotiledóneas y Monocotiledóneas.

29 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
especies, Fabáceas (=Leguminosas) con
18.000 especies, y Poáceas (=Gramíneas)
con casi 10.000 especies. Aunque no son
tan importantes para el hombre ni
dominantes en el paisaje, las Orquideáceas,
con casi 20.000 especies, constituyen la
segunda familia más numerosa de todas, y
es muy relevante en los ecosistemas
selváticos tropicales.
Ciclo biológico
Las Angiospermas, como en todas las
Espermatofitas (Plantas con Semillas),
presentan alternancia de dos generaciones
multicelulares enmascaradas, ya que el
gametofito se desarrolla por completo
dentro de las estructuras del esporofito, el
cual se encuentra organizado en un "cormo"
o sea que presenta raíz y vástago (tallo +
hojas), que incluyen un sistema de
conducción desarrollado. Al igual que las
Gimnospermas, poseen un ciclo de vida
heterosporado y los gametofitos se
desarrollan en forma endospórica (esto
quiere decir que el gametofito femenino se
desarrolla dentro de la macrospora y el
gametofito masculino en el interior de las
microsporas). Los esporangios se
encuentran siempre en antófilos fértiles
(=esporofilos, es decir estambres y
carpelos), los cuales se encuentran
formando parte de una estructura llamada
flor.
Los carpelos o megasporofilos son hojas
fértiles cerradas que constituyen el gineceo
y se encuentran diferenciados en una
superficie preparada para recibir al grano de
polen llamada estigma, un porción alargada
(estilo) y una cavidad que alberga a los
óvulos llamada ovario. Los óvulos o
megasporangios están constituidos por los
tegumentos y un tejido fértil llamado nucelo
en el cual se diferencia la célula madre de la
megaspora que por meiosis da lugar a
cuatro megasporas, tres de las cuales
degeneran persistiendo solo una. Esta
megaspora se divide mitóticamente
originando ocho núcleos. Una vez formados
los tabiques quedan diferenciadas siete
células, una de ellas, la central, es
binucleada. Esta estructura compuesta por
siete células recibe el nombre de saco
embrionario y constituye el gametofito
femenino o megagametofito.
Por lo general, los estambres o
microsporofilos poseen en su porción fértil
o antera, con cuatro microsporangios o
sacos polínicos. En el tejido fértil o
arquesporio se diferencian las células
madres de las microsporas las cuales por
meiosis originan cuatro microsporas o
"granos de polen". El gametofito masculino
o microgametofito está reducido a dos
células, una vegetativa que origina el tubo
polínico y una generativa que da lugar a los
núcleos espermáticos o gametas
masculinas. Una vez depositado el grano de
polen en el estigma el tubo polínico
comienza a germinar a través de uno de los
poros. El crecimiento del mismo está
dirigido por la célula vegetativa y la
expansión es a partir de la intina. El tubo
polínico llega hasta la micrópila del óvulo
donde vuelca su contenido. En las
Angiospermas se produce el fenómeno de
doble fecundación: uno de los dos núcleos
espermáticos que libera el grano de polen
fecunda a la ovocélula o gameta femenina
(produciendo el cigoto o primera célula
esporofítica) y el otro a los núcleos polares
de célula diploide del saco embrionario,
originando un tejido 3n (triploide) de
reserva llamado endosperma.
El cigoto, por mitosis, se transforma en un
embrión, que al madurar entra en estado de
latencia rodeado de restos de nucelo y/o
endosperma y los tegumentos del óvulo,
que en conjunto forman la semilla. La
semilla madura es liberada convirtiéndose
en la unidad de dispersión y en condiciones
adecuadas germina. La estructura particular
de las flores de las Angiospermas no es la
única diferencia que poseen con el resto de
las Espermatofitas, otros caracteres
morfológicos distintivos son: la reducción
del gametofito femenino a sólo unas pocas
células y la doble fecundación con la
formación de un tejido nutritivo
característico 3n llamado endosperma, al
tiempo que se fecunda a la ovocélula con la
consiguiente formación del cigoto.

30 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Ciclo biológico de una Angiosperma. Tomado de Wikipedia.
Angiosperma

31 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
4.1. TEJIDOS Introducción
El cuerpo de las plantas superiores consta
de una gran variedad de células que difieren
en tamaño, forma y otros caracteres.
Normalmente, en el cuerpo del vegetal hay
numerosas células con la misma función
reunidas en complejos llamados tejidos.
Los tejidos meristemáticos son
aquellos que conservan durante toda su vida
la capacidad de dividirse y formar células y
pueden subdividirse en meristemas
primarios, que conservan su carácter
mersitemático desde su desarrollo
embrionario; los meristemas secundarios
en cambio, se originan a partir de células de
tejidos adultos que “recuperan” su
capacidad de división.
Los tejidos definitivos son los que
constituyen la mayor parte de un vegetal
adulto y se pueden distinguir según su
función en: parénquimas
(=fundamentales), de aislación, de sostén,
de conducción, de absorción y de
secreción.
Se denominan simples a los tejidos
constituidos por células similares mientras
que los que presentan dos o más tipos de
células son los llamados tejidos complejos.
TEJIDOS MERISTEMÁTICOS
Están formados por células jóvenes que
persisten en el cuerpo de las plantas adultas
y que conservan su capacidad de división
mitótica. Por lo general poseen paredes
delgadas, con vacuolas pequeñas,
numerosas mitocondrias, plástidos en
estado proplastidial y núcleo grande.
Según su origen, los meristemas pueden
ser: meristemas primarios
(=promeristemas), que son los que
provienen siempre de células embrionarias
(meristemas apicales, responsables del
crecimiento en longitud de los distintos
órganos) mientras que los meristemas
secundariosson los que provienen de
células adultas que por un proceso llamado
de “dediferenciación” adquieren
características meristemáticas (meristemas
laterales: responsables del aumento del
diámetro de los distintos órganos).
Según la posición en el cuerpo de la planta,
se pueden distinguir a los meristemas
apicales: en los ápices de tallos y raíces
(responsables del crecimiento en longitud);
los meristemas laterales: ubicados en
forma paralela a la superficie del órgano en
el que se presentan, representados por el
cámbium, que produce los tejidos de
conducción y el felógeno, que produce un
tejido de aislación, la peridermis. Los
meristemas intercalares, se ubican entre
tejidos bien diferenciados y el ejemplo más
común es el que se ubica en la base del
nudo de los tallos de las cañas (Gramíneas)
y que produce el crecimiento del entrenudo.
Las células meristemáticas pueden dividirse
en distintos sentidos: cuando la división es
perpendicular a la superficie del meristema
se produce un crecimiento en superficie,
mientras que si las divisiones son paralelas,
el crecimiento será en volumen,
aumentando el espesor del cuerpo. En el
caso que un grupo de células aisladas posea
la capacidad de dividirse reciben la
denominación de meristemoides.
TEJIDOS ADULTOS
Todos los tejidos adultos derivan de
meristemas, pero a diferencia de éstos, han
perdido su capacidad de división y
adquieren formas y características propias
de la función de cada tejido.
3.1. Parénquimas: las masas de células con
relativamente poca especialización en
estructura o función, se denominan
parénquimas fundamentales, como por

32 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
ejemplo, la porción central de algunos los
tallos jóvenes (médula). En el caso de estar
especializados en la acumulación de
sustancias de reserva (como carbohidratos o
lípidos), reciben el nombre de parénquimas
reservantes, comunes en raíces, frutos y
algunos tallos. Probablemente los tejidos
parenquimáticos con mayor grado de
especialización son los clorénquimas o
parénquimas clorofilianos que constituyen
la parte interna de las hojas (y tallos jóvenes
o modificados) donde se realiza el proceso
de fotosíntesis.
Tejidos de aislación:
Epidermis: la superficie de todas las partes
“blandas” de las plantas está formada, en
general, por una sola capa de células las
capas exteriores de células denominada
epidermis. Esta epidermis está
especializada para evitar la pérdida
excesiva de agua de los tejidos subyacentes,
lo que se logra mediante una modificación
de la composición química de las paredes
externas de las células que la constituyen,
ya que éstas son impregnadas por una
sustancia segregada por las mismas células
llamada cutina, un material ceroso e
impermeable que constituye una deposición
delgada y compacta: la cutícula. La
cutícula no es continua, ya que sobre todo
en las hojas y en tallos jóvenes, presenta
poros o estomas que permiten el
intercambio gaseoso con el medio
ambiente. En algunos casos, la epidermis
posee más de un estrato de células,
recibiendo el nombre de epidermis
pluriestratificada.
Hipodermis: tiene el aspecto y cumple la
misma función que una epidermis
pluriestratificada, pero es de un origen
distinto a la epidermis propiamente dicha.
Endodermis: es un tejido formado una capa
de células alargadas que limitan tejidos
internos, característica de las raíces de las
plantas superiores (excepcionalmente en
algunos tallos y hojas), entre la corteza y
los tejidos vasculares (cilindro central), en
las que una sustancia impermeable parecida
a la suberina, llamada endodermina, se
deposita en determinadas zonas de las
paredes celulares formando una banda
continua denominada banda de Caspary,
cuya función es controlar la difusión
ilimitada de agua y solutos. Más adelante,
toda la célula se recubre interiormente de
endodermina, salvo algunas células que
permanecen sin deposición, son permeables
y reciben el nombre de células de paso.
Peridermis:cuando los tallos jóvenes crecen
considerablemente en grosor, la epidermis
es reemplazada por un conjunto de tejidos
más resistentes: la peridermis, que se
origina por la acción de un meristema
lateral, el felógeno, que produce felodermis
(parenquimática) hacia el interior y súber o
corcho (células que se suberinizan y
mueren) hacia la parte externa. Los estomas
de la epidermis son reemplazados en la
peridermis por lenticelas, cuya función es
también el intercambio gaseoso, pero con
una estructura más compleja.
Tejidos de sostén
Colénquima: es un tejido formado por
células siempre vivas, que puede
considerarse también como un parénquima
especializado en la función de sostén. Las
células de colénquima poseen las paredes
celulares muy engrosadas con celulosa y
sustancias pécticas. Estos engrosamientos
no se producen siempre en forma uniforme:
si se limitan a los ángulos de las células, el
colénquima recibe la denominación de
“angular”; si la deposición extra de
celulosa se produce en las paredes laterales,
será “laminar” y por último, el
colénquima “lagunar” se produce cuando
el tejido presenta espacios intercelulares
grandes y los engrosamientos se producen
en las paredes que limitan esos espacios.
Pese a este engrosamiento, son capaces de
sufrir dilatación y crecer, por lo que se
encuentran sobre todo en las partes jóvenes
de la planta que están en activo crecimiento.
Esclerénquima: es un tejido formado por
células siempre muertas y a diferencia del
colénquima, las paredes secundarias de las
células esclerenquimáticas tienen
engrosamientos más o menos uniformes y
están impregnadas de lignina, una
sustancia que les confiere gran resistencia
mecánica (especialmente a la presión). Los
engrosamientos ocupan la mayor parte del
lumen celular, por lo que una vez finalizada
la síntesis de las sustancias que forman la

33 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
pared, el protoplasto muere. Se distinguen
dos tipos de células esclerenquimáticas: las
fibras, alargadas con los extremos aguzados
y las esclereidas o células pétreas, más o
menos isodiamétricas de formas variadas.
Tejidos de conducción
El xilema y el floema se encuentran
relacionados espacialmente y constituyen el
sistema vascular de todo el cuerpo del
vegetal. Son tejidos complejos cuyas
células han alcanzado una importante
especialización.
Xilema: el xilema es el tejido responsable
de transportar el agua y las sales que
absorben las raíces hacia todos los demás
órganos de la planta. Este movimiento,
principalmente ascendente recibe el nombre
de “acrópeto”. El xilema está constituido
por células parenquimáticas (parénquima
xilemático), por fibras esclerenquimáticas
(=fibras xilemáticas) y por traqueidas y
elementos de tráqueas (= elementos de
vasos).
Traqueidas: células que mueren a la
madurez, alargadas, con los extremos
agudos que sirven para la conducción de
agua y solutos. Debido a su función
conductora, sus paredes son ricas en
punteaduras (perforaciones),
principalmente en las paredes terminales
oblicuas, para facilitar el paso en dirección
longitudinal. También funcionan como
elementos de sostén ya que las paredes
secundarias están engrosadas y
lignificadas.
Elementos de tráqueas: son células sin
citoplasma a la madurez, menos elongadas
pero de mayor lumen que las traqueidas,
con las paredes secundarias también
engrosadas y lignificadas. A diferencia de
éstas, los elementos de tráquea presentan
perforadas las paredes distales y encajan
unos con otros formando largos tubos
continuos llamados tráqueas o vasos. En
las tráqueas el agua se mueve libremente y
solo debe atravesar la primera y última
pared del primer y último elemento
respectivamente. Para evitar la compresión
de los vasos debido a la fuerza de succión
de la columna de agua que sube por ellos,
están reforzados por engrosamientos en la
pared secundaria. Al mismo tiempo, como
debe conservarse el intercambio de agua
con las células contiguas, los
engrosamientos son parciales y tienen
forma de espiral, de anillos, de redes y en
los vasos punteados, prácticamente toda la
pared está engrosada y sólo las punteaduras
son delgadas.
Floema: es el tejido responsable de
transportar los productos de la fotosíntesis
desde las hojas hacia el resto del cuerpo del
vegetal. Del mismo modo que en el xilema,
en la constitución del floema intervienen
células parenquimáticas y fibras. Las
células que caracterizan al floema son los
tubos cribosos: células alargadas que en las
paredes de los extremos (en algunos grupos
de plantas en las paredes laterales también),
presentan zonas perforadas, los campos
cribosos; cuando éstos se unen forman una
gran región perforada llamada placa
cribosa. Tanto los campos como las placas
cribosas están tapizados por calosa, un
polisacárido estructural que es el que obtura
los poros cuando el tubo criboso deja de ser
funcional. A través de las placas, pasan
bandas de citoplasma de una célula a otra,
obteniéndose así una eficiente
comunicación. Las paredes de los tubos
cribosos están muy poco engrosadas. En
general se consideran carentes de núcleo
(en algunos casos sí poseen) y adyacentes a
ellos y de menor tamaño, se disponen las
células llamadas acompañantes o anexas
en Angiospermas, que provienen de la
misma “célula madre” del tubo criboso y sí
presentan núcleo. En el caso de las
Gimnospermas, estas células reciben el
nombre de albuminíferas. Generalmente
los tubos cribosos están activos durante un
período vegetativo y al morir el elemento
de tubo criboso, muere también la célula
acompañante.
Tejidos de absorción
Rizodermis: recibe este nombre la
epidermis que recubre la raíz, y que a
diferencia de la epidermis de los órganos
aéreos que está especializada en evitar la
pérdida excesiva de agua, su función
principal es la absorción de agua y sales del
suelo, por lo que las células de este tejido
carecen de cutícula. La zona denominada de
los “pelos absorbentes” presenta las células
de la rizodermis transformadas en pelos,

34 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
que al aumentar la superficie son muy
eficientes en la absorción. Algunas veces
hay una diferenciación morfológica de las
células que originan los pelos y reciben el
nombre de tricoblastos.
Velamen: es un tejido constituido por
células muertas que rodean la raíz de las
orquídeas epífitas y de otro grupo de
plantas también epífitas (aráceas).
Absorben vapor de agua (o agua) y también
nitrógeno del aire como si fueran un papel
secante y los transfieren a las partes más
internas del órgano.
Tejidos de secreción
Laticíferos: se designan así a tejidos
formados por células o series de células que
contienen un líquido llamado látex y que
constituyen sistemas que atraviesan
diferentes órganos. El látex está formado
por una emulsión de proteínas, azúcares,
gomas, resinas, alcaloides, aceites
esenciales y otras sustancias. De estas
sustancias, los hidrocarburos relacionados
con el caucho y las resinas, constituyen el
látex de muchas plantas. Generalmente
están asociados al floema, pero no es
excluyente.
Conductos resiníferos: algunas
Gimnospermas (pinos, tuyas, etc.)
presentan conductos que se originan como
espacios intercelulares mediante la
separación de células parenquimáticas
productoras de resinas. Los conductos
quedan tapizados por estas células,
responsables de la síntesis de resinas. Su
presencia está asociada muchas veces a
situaciones traumáticas y en relación directa
con el xilema secundario de los tallos y
también, formando parte del parénquima
clorofiliano las hojas.

35 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
4.3. HOJA Características y Funciones
Las hojas son los órganos fotosintéticos de
la mayoría de las plantas vasculares. La
función fundamental de las hojas es la
fotosíntesis y a ésta va asociada
indefectiblemente la transpiración de
vapor de agua, proceso que es a la vez una
necesidad y un problema para las plantas.
Para cumplir con la función fundamental las
hojas comunmente son aplanadas, con una
gran superficie en relación al volumen y
con simetría dorsiventral. Entre las
funciones adicionales están el almacenaje
de agua (plantas suculentas) o sustancias
alimenticias (hojas almacenadoras de los
bulbos tunicados); protección, ya sea
recubriendo otros órganos (catáfilos) o
como espinas; soporte, el caso de las hojas
modificadas en zarcillos.
Las hojas poseen dimensiones muy
variadas, que pueden ir de pocos milímetros
a varios metros. La hojas más grandes se
encuentran en las Palmeras. La hojas de
grandes dimensiones presentan problemas
mecánicos para soportar el viento, por ello
muchas veces están divididas. En las
Palmeras esto se logra por medio de bandas
de necrosis espontánea que se producen
durante el desarrollo temprano de la hoja.
Las hojas simples enteras más grandes del
reino vegetal se presentan en la familia de
las Aráceas. Algunas de las mas pequeñas
en las Lemnáceas, una familia
estrechamente emparentada con la anterior.
Figura 4.3.1. Xanthosoma sp. (Araceae)
Figura 4.3.2. Spirodella intermedia
(Lemnaceae) está constituida por una hoja
flotante de 1-3 mm, un tallo reducido a pocas
células y una raíz de pocos cm. Esta especie se
incluía en la familia Lemnaceae.
El follaje de las plantas que cubre una
determinada porción de suelo puede llegar a
tener una superficie varias veces mayor que
aquella. Esta proporciónse conoce como
índice de área foliar, que representa
basicamente los metros cuadrados de hojas
que se encuentran por encima de un metro
cuadrado de suelo. Este suele ser menor que
1 (uno) en los desiertos y ambientes áridos,
alcanzar valores máximos de 7 en pastizales
sin pastoreo y valores algo mayores aún en
las selvas.
El término hoja se aplica en sentido amplio
tanto a plantas con semillas, pteridofitas y
briofitas. Sin embargo, los órganos
aplanados y fotosintéticos de las briofítas
no son estrictamente homólogos a los de los
otros grupos y algunos autores los
denominan filidios o filoides. Por otro lado
en algunas pteridofitas pueden distinguirse
hojas muy simples con una sola nervadura
que se denominan micrófilos, mientras que
otras poseen hojas estructuralmente
complejas con un sistema desarrollado de
nervaduras y reciben el nombre de
macrófilos o megáfilos. Las hojas de los
helechos reciben con frecuencia el nombre
de frondes.
Secuencia foliar durante la ontogenia
Durante las fases de desarrollo de una
planta las hojas presentan transformaciones
paulatinas tanto desde el punto de vista

36 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
morfológico como funcional. Las primeras
hojas son las embrionales y reciben el
nombre de cotiledones, frecuentemente
especializadas en el almacenaje de
sustancias nutritivas. A continuación,
usualmente mientras el tallo tiene
crecimiento subterráneo, aparecen los
catáfilos, hojas escamosas y muchas veces
sin pigmentos que cumplen funciones de
protección. Luego, las hojas fotosintéticas
reciben el nombre de nomófilos. En
muchos casos los nomófilos que aparecen a
lo largo de un tallo van ganando en
complejidad estructural. Las hojas
asociadas a flores o inflorescencias reciben
el nombre hipsófilos o brácteas, cumplen
función de atracción a polinizadores. Por
último las hojas altamente modificadas que
forman las distintas piezas florales, y que
reciben el nombre de antófilos, también
sufren transformaciones y aumento de
complejidad.
Figura 4.3.3. Esquema de un eje caulinar con
los distintos tipos de hojas.
Filotaxis
Las hojas siempre se insertan en los nudos
del tallo. Filotaxis es la disposición de las
hojas en el tallo. Si existe una sola hoja en
cada nudo se habla de filotaxis esparcida u
hojas alternas. Si aparecen dos hojas en
cada uno se habla de hojas opuestas. Si
aparecen más de dos hojas en cada nudo se
habla de filotaxis verticilada.
Figura 2. Filotaxis.
Partes de la hoja
La morfología foliar se describe
considerando una amplia gama de aspectos
que pueden tener importancia para
caracterizar especies u otros niveles
taxonómicos. Sin embargo, en muchos
casos es esencialmente un complemento en
la clasificación, ya que especies
taxonómicamente muy distantes pueden
tener hojas muy parecidas.
Un nomófilo completo tiene una serie de
partes: la base foliar, es decir la zona de
inserción en el tallo. En varias familias de
plantas las hojas tienen la base formando
una vaina que envuelve al tallo. A partir de
la base foliar existe una prolongación
normalmente angosta que recibe el nombre
de pecíolo. El pecíolo conecta la base o
vaina con la lámina o limbo, que es la
porción ensanchada y especializada en el
proceso de fotosíntesis. La lámina tiene a su
vez una base, la lámina propiamente dicha y
un ápice que tienen formas características.
Figura 4.3.4. Partes de la hoja.
En algunos casos las hojas que se insertan
en un mismo nudo o en una posición

37 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
cercana en el tallo tienen dimensiones
marcadamente diferentes, fenómeno que
recibe el nombre de anisofilia. Por su parte
las hojas pueden ser marcadamente
diferentes si están sometidas a condiciones
ambientales diferentes, como es caso de las
hojas aéreas y las acuáticas, sumergidas, de
algunas plantas. Se habla en este caso de
heterofilia.
Los tejidos de conducción de la hoja se
disponen formando filamentos que reciben
el nombre de nervaduras o venas. La
disposición de estas nervaduras se conoce
como nerviación o venación. La
nerviación tiene formas características,
siendo usualmente retinervada en las
dicotiledóneas y paralelinervada en las
monocotiledóneas. La nerviación
dicotómica es una estructura primitiva que
se encuentra en varios helechos y en
Ginkgo biloba (Gimnosperma).
Hojas simples y compuestas
La lámina puede estar constituída por una
sola pieza o estar compuesta por varias
unidades o folíolos. Estos pueden estar a su
vez subdivididos en foliólulos. Las hojas
compuestas pueden tener los folíolos
distribuídos en forma palmada o pinnada.
Un criterio muy importante para distinguir
una hoja con varios folíolos de una rama
con varias hojas está dado por la posición
relativa de las yemas laterales.
Figura 4.3.5.Hojas simples y compuestas
Figura 4.3.6.Tipos de ápice foliar
Figura 4.3.7. Tipos de base foliar
Figura 4.3.8.Formas comunes de hoja.
Apéndices foliares
En la base de las hojas, en muchos grupos
de plantas, están presentes dos pequeñas
piezas laterales llamadas estípulas. En
casos excepcionales éstas poseen mayor
tamaño que la lámina misma. En la familia
de las Poligonáceas las estípulas se
encuentran soldadas formando una cubierta
que envuelve al tallo, denominada ocrea.
En las Gramíneas y otras Monocotiledóneas
las hojas poseen una vaina con dos
apéndices laterales, denominados

38 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
aurículas, en el punto de unión con la
lámina. Estas aurículas tienen formas
carácterísticas y permiten a veces
identificar especies en estado vegetativo.
Esto es particularmente usado para
identificar los llamados “cereales
invernales” (trigo, avena, cebada, centeno)
antes de floración. En esa misma zona, en la
inflexión entre la vaina y la lámina existe
un apéndice membranoso que recibe el
nómbre de lígula, que también tiene gran
utilidad taxonómica en las Gramíneas. En la
base del pecíolo o en la unión del pecíolo
con la lámina existe en algunos grupos un
engrosamiento que recibe el nombre de
pulvínulo. En ciertas familias
(Leguminosas) el pulvínulo puede ser capaz
de realizar movimientos por cambios de
turgencia, ya sea en distintos momentos del
día o como respuesta a estímulos tactiles.
Estos reciben el nombre de pulvínulo
motor.
Prefoliación, Vernación y Ptyxis
Se llama vernación o prefoliación a la
posición y plegamiento que adopta la hoja
en la
yema.
Figura 4.3.9. Tipos de vernación. A: reclinada o
inflexa, con el ápice curvado hacia la base; B:
conduplicada, es decir plegada a lo largo de la
nervadura central; C: Plegada, a lo largo de las
venas princiaples como un abanico; D:
Circinada, enrollada sobre si misma desde el
ápice a la base; E: convoluta, enrollada
longitudinalmente sobre si misma; F: involuta,
con los bordes enrollados sobre la cara superior;
G: revoluta, con los bordes enrollados sobre la
cara inferior.
En los textos en inglés lo descripto aquí
como vernación recibe el nombre de ptyxis,
mientras que se reserva el término
vernation a la posición relativa de las
distintas piezas foliares, por ejemplo
imbricadas, cuando se hallan parcialmente
superpuestas o valvadas cuando están en
contacto solamente por sus márgenes (ver
también prefloración en el capítulo de
flor).
Profilo
La primera o dos primeras hojas que surgen
en una yema lateral reciben el nombre de
profilo y suelen ser mofológicamente algo
diferentes de las siguientes. En las
dicotiledóneas suele haber dos profilos,
mientras que en las monocotiledóneas se
presenta uno solo con dos nervaduras
medias que se interpreta como resultante de
la soldadura de dos piezas en el curso de la
evolución.
Modificaciones adaptativas de la hoja
Como se vió en raíz y tallo, la hoja también
ha sufrido modificaciones adaptativas para
cumplir funciones especiales.
Zarcillos. Son formaciones filiformes que
permiten fijarse a distintos soportes y crecer
sustentados por ellos. La lámina de las
hojas transformadas pueden serlo total o
parcialmente, dejando algunos folíolos
fotosintéticos.
Espinas. Son hojas transformadas en
estructuras punzantes que actúan como
repelentes de herbívoros. En algunos casos
las estípulas están transformadas en espinas,
mientras que la lámina conserva su función
normal. En algunas Cactáceas (ej.: Opuntia
ficus-indica) numerosas hojas de una yema
se transforman en pequeñas estructuras
punzantes (“janas”), que se desprenden
cuando el filocladio que las posee es
disturbado. Estas hojitas modificadas
punzantes reciben el nombre de gloquídios.
Hojas almacenadoras. Las hojas pueden
cumplir funciones de reserva, como en los
bulbos tunicados (ej.: Allium cepa, la
cebolla). Estos están formados en su mayor
parte por la base engrosada de las hojas, en
la cuales la porción fotosintética superior
muere a la madurez.
Hojas suculentas. La especialización
puede ser para almacenar agua, como
sucede en algunas familias (ej.:
Portulacáceas, Crasuláceas) adaptadas para
la vida en condiciones de aridez.
Pérulas. Las yemas invernales de los
árboles de ambientes templados y fríos
suelen estar cubiertas por hojas muy poco

39 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
expandidas resistentes a la desecación y al
ataque de insectos, que se desprenden
cuando la yema comienza su actividad
primaveral.
Filodio. Los filodios son pecíolos
transformados en estructuras
fotosintetizadoras, usualmente por pérdida
del limbo en el curso de la evolución.
Hojas carnívoras. Existen plantas
adaptadas a ambientes con bajo contenido
de nitrógeno en el sustrato, que compensan
su deficiencia capturando pequeños
animales. La captura la realizan hojas
modificadas con distintos mecanismos de
cierre o glándulas adhesivas que
inmovilizan a la presa. La familia
Droseráceas es una de las que posee
representantes carnívoros.
Espatas. Son hojas modificadas asociadas a
una inflorescencia. Por ejemplo la vistosa
hoja roja de Anthurium o la blanca de
Zanthedeschia aethiopica son espatas, que
rodean a una inflorescencia carnosa llamada
espádice.
Estructura anatómica de la lámina
La estructura de la lámina está orientada a
dos necesidades principales, la realización
de la fotosíntesis y la regulación de la
pérdida de agua. Está constituida por tres
tejidos principales: un tejido superficial
protector (epidermis), un tejido
fotosintético (mesófilo) y tejidos de
conducción y sostén de la estructura (haces
vasculares).
Epidermis
La epidermis desempeña dos funciones
contrapuestas: por un lado limita la pérdida
de agua al exterior protegiendo de la
desecación y por el otro permite el
intercambio gaseoso con el ambiente. Para
cumplir estas funciones la epidermis forma
una película altamente impermeable que
envuelve la hoja, formada por una sola capa
de células recubiertas en su cara externa por
una sustancia denominada cutina, sobre la
que pueden depositarse ceras. Esta película
está interrumpida por gran cantidad de
poros, denominados estomas, que permiten
el pasaje de gases, de una manera
controlada. Los estomas trabajan para
optimizar la absorción de CO2, que se
encuentra en concentraciones relativas muy
bajas en la atmósfera (ver tabla 4.3.2) y
moderar la salida de agua, que se encuentra
en estado líquido en el interior de la hoja y
tiende a pasar al estado gaseoso presente en
la atmósfera, en la mayor parte de las
condiciones. La epidermis frecuentemente
presenta pelos o tricomas, que cumplen
funciones variadas de protección,
principalmente contra insectos y ácaros
fitófagos.
Figura 4.3.10. Típicos estomas de dicotiledónea
vistos con un microscopio electrónico de
barrido.
Figura 4.3.11. Epidermis con alta densidad de
tricomas eglandulares (alargados, no producen
secreciones, tiene acción mecánica) y
glandulares (cortos, con una cabeza esférica que
acumula secreciones anti-herbívoro).
Control estomático
Las plantas se encuentran ante un conflicto
existencial: abrir los estomas para poder
absorber CO2 o cerrarlos para evitar la
pérdida excesiva de agua. Los estomas
poseen un complicado sistema de control de
apertura y cierre para resolver este dilema y
hacer eficiente al máximo el proceso. Esto
es, ganar la mayor cantidad posible de CO2
en relación al vapor de agua perdido. El
aparato estomático responde a una serie de

40 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
estímulos internos y externos sintetizados
en la tabla 4.3.1. El mecanismo de apertura
y cierre está controlado por la salida o
entrada de iones K+, que altamente
hidratados, determinan la mayor o menor
turgencia de las células oclusivas.
Factor Estado Respuesta
Condición
hídrica
Buena Apertura
Deficitaria Cierre
CO2 interno de
la hoja
Elevado Cierre
Bajo Apertura
Ácido Absísico Elevado Cierre
Bajo Apertura
Luz Intensa Apertura
Pobre u oscuridad Cierre
Tabla 4.3.1.Principales factores que regulan la
apertura y cierre de los estomas.
Mesófilo
El tejido fundamental que ocupa el espacio
entre ambas caras epidérmicas recibe el
nombre de mesófilo y es el responsable de
llevar a cabo toda o la mayor parte de la
actividad fotosintética de la planta. Este
tejido recibe el nombre de parénquima
clorofiliano, que puede estar
homogeneamente distribuído (por ejemplo
en las hojas de Gramíneas) o estar
diferenciado en dos tipos, como parénquima
lagunar o esponjoso y parénquima en
empalizada. Ambos poseen importantes
espacios intercelulares que facilitan el
intercambio gaseoso, esencialmente ingreso
de CO2 y la liberacion de O2 y vapor de
H2O a las células fotosintéticas. El
parénquima en empalizada suele concentrar
el 80% de los cloroplastos de la hoja,
mientras que el lagunoso posee el 20%
restante.
Haces vasculares
El tercer elemento que componen las hojas
es la red de haces vasculares que
suministran agua al sistema fotosintético,
transportan las sustancias elaboradas al
resto de la planta y dan soporte estructural a
la lámina foliar. Los haces vasculares son
generalmente colaterales cerrados, con
floema del lado del envés y el xilema del
lado del haz. Generalmente asociados a los
haces vasculares, se encuentran tejidos de
sostén.
Absición
La mayoría de las hojas de los helechos y
angiospermas herbáceas cuando senescen se
marchitan en el tallo y caen por
descomposición o fuerzas externas. En
cambio la mayoría de las hojas de
gimnospermas y angiospermas leñosas
generalmente caen por la formación de una
banda de absición en la base del pecíolo.
En esta zona existe una proporción alta de
células parenquimáticas de paredes
delgadas y poca densidad de tejido vascular
o de sostén. En el proceso de absición se
forman dos capas, una capa protectora de
células suberificadas en contacto con el
tallo y otra capa de separación formada
por células cuyas paredes sufren un proceso
de degradación de la laminilla media.
El proceso de absición puede
desencadenarse por distintos factores
ambientales como falta de agua, bajas
temperaturas o días cortos.

41 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Figura 4.3.12. Corte transversal de una hoja de dicotilendónea. Se observa dos tipos de
parénquima clorofiliano, el parenquima en empalizada, ubicado en la parte superior y el
parénquima esponjoso, con grandes lagunas. Puede verse la epidermis uniestratificada, con
células con citoplasma menos denso que las del parénquima fotosintético.
Figura 4.3.13. Corte transversal de una hoja de Nerium oleander, una especie adaptada a
ambientes semiáridos. Posee una epidermis pluriestratificada y con una cutícula gruesa. Los
estomas se encuentran en criptas de la epidermis inferior, con pelos. Estas criptas poseen niveles
de humedad más elevados que permiten mantener abiertos los estomas, reduciendo la
transpiración.

42 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Figura 4.3.14. Corte transversal de una hoja de una gramínea, caña de azúcar (Saccharum
officinarum). Las gramíneas poseen un solo tipo de parénquima fotosintético (homogéneo). En
la epidermis superior e inmediatamente por debajo existen grandes células denominadas
buliformes. Estas células pierden mucho volumen con la deshidración, provocando la curvatura
de la hoja sobre la cara superior, lo que reduce la superficie de evaporación.
Figura 4.3.15. Corte transversal de una hoja acicular de pino. Estas hojas están bien adaptadas
al estrés hídrico, con estomas deprimidos con respecto a la epidermis, que posee una cutícula
gruesa y una hipodermis esclerificada. El parénquima clorofiliano es homogéneo. El haz
vascular se encuentra rodeado por una capa endodérmica. Se observan además conductos
resiníferos.

43 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
4.4. TALLO
Caracterísicas y funciones
El tallo es el órgano que sirve de nexo entre
las hojas, es decir los órganos
fotosintetizadores y el sistema radicular, o sea
las estructuras de absorción y fijación al
sustrato.
El tallo cumple entonces funciones de soporte
de las hojas, constituyendo una estructura
fuerte y flexible que permite ubicar las hojas
de modo que fotosinteticen de manera óptima;
soporte de las flores, ubicando las flores en
una posición adecuada para la polinización;
soporte de los frutos, sosteniendo el peso de
los frutos y exponerlos para su dispersión. Por
otro lado cumple una función de conducción
de agua y minerales, ya que los tejidos del
xilema de la raíz se continúan en el tallo y a
través de ellos se distribuyen agua y solutos en
el follaje; conducción de sustancias
nutritivas a través del floema hacia toda la
planta, desde las hojas donde se producen los
azúcares. Con frecuencia es el órgano más
voluminoso de la planta y cumple funciones de
almacenamiento de sustancias nutritivas
(azúcares simples, almidón) o agua. Por último
los tallos pueden cumplir funciones de
protección, modificados en espinas.
Dimensiones y longevidad
El tamaño de los tallos es muy variable según
las especies y las condiciones de crecimiento.
Como ejemplo de tallos mínimos puede
mencionarse a especies de la familia de las
Lemnáceas (lentejas de agua), de los
génerosSpirodela (ver Figura 4.3.2),Lemna y
Wolfia, en los cuales el tallo practicamente ha
desaparecido,limitándose a unas pocas células,
y la planta entera tiene 1-3 mm. En el otro
extremo se encuentran los árboles, en los cuales
el tallo es con frecuencia el órgano más
voluminoso. Los árboles más grandes son las
sequoias o redwoods (Sequoiadendron
giganteum, Figura 3.3.4), que alcanzan varias
decenas de metros de altura (117 m) y
diámetros de hasta 7 m. También dentro de las
gimnospermas se encuentran los árboles mas
longevos, el bristlecone pine (Pinus longaeva)
4900 años, en América del Norte y el alerce
(Fitzroya cupresoides), 3600 años, en los
Andes de la Patagonia de Chile y Argentina.Las
angiospermas más altas y grandes pertenecen al
género de los eucaliptos. Eucalyptus jacksonii,
en el sudoeste de Australia alcanza más de 100
m de alto y alrededor de 20 m de perímetro en
la base. Un ejemplar de este género de más de
120m, talado en Australia hace décadas, fue el
árbol más alto conocido en tiempos históricos.
Los tallos más grandes en Sudamérica
pertenecen probablemente alFitzroya
cupresoides. En el NOA los tallos más grandes
pertenecen al laurel (Cinammomum porphyria)
y al cedro "tucumano" (Cedrela lilloi). El
cedro más grande que se conoce, un ejemplar
de C. lilloi encontrado en Osma, provincia de
Salta, tiene 3 m a la altura del pecho (1.40 m) y
una altura de 25 m (Fig. 4.4.1). Es probable que
en el pasado no muy lejano halla habido árboles
más grandes aún. En relatos se menciona que la
selva entre Tucumán y la sierra de San Javier
"tenía árboles tan grandes que se necesitaban 13
hombres tomados de la mano para rodearlos".
El horco molle (Blepharocalyx salicifolia = B.
gigantea) también presenta ejemplares de gran
tamaño
Figura 4.4.1. Cedrela lilloi (Meliaceae) en la
localidad de Osma, Salta. El ejemplar viviente de
cedro más grande conocido en la actualidad.
(Fotografía gentileza Roberto Neumann).
Una proporción muy importante del carbono
de la tierra se encuentra en los tallos de
vegetales, en los bosques y selvas. Otra
proporción muy importante se encuentra en los
tallos de vegetales muertos hace millones de

44 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
años, formando los depósitos de carbón de
piedra. La liberación de este carbono a la
atmósfera por combustión es una de las causas
del incremento en la concentración en la
atmósfera. Hay quienes proponen la necesidad
de plantaciones masivas de arboles para
"secuestrar" nuevamente un parte sustancial de
este carbono y reducir su concentración
atmosférica. Estos proyectos cuentan con el
apoyo de ciertos sectores, con el argumento de
que permitirían en países del tercer mundo un
desarrollo forestal importante, que se podría
asociar también a la creación de nuevas áreas
protegidas. Otros en cambio critican la idea,
argumentando que la reducción de carbono
atmosférico debería centrarse en la reducción
de emisiones por parte de los países
desarrollados cuya tasa de emisión per cápita
supera holgadamente a la de los países en
desarrollo.
Morfología
Los tallos son órganos con simetría radial.
Comunmente es posible diferenciar en ellos dos
elementos: entrenudos (o internodios) y
nudos. El nudo es la porción del tallo donde se
insertan las hojas. En esa región algunos haces
vasculares, que en el entrenudo ocupan una
posición interna, se dirigen hacia el exterior e
inervan la hoja correspondiente. En el extremo
del tallo existe usualmente una yema apical. En
ella las distintas hojas son todavía pequeñas
(primordios foliares) y las distancia entre éllas
es mínima. Por ello, allí los nudos y entrenudos
son difíciles de distinguir. El alargamiento del
tallo da origen a los entrenudos. En la axila de
cada hoja existe comunmente una yema, que es
un primorido de vástago, es decir un meristema
de crecimiento que puede originar una
ramificación lateral.
La mayoría de las yemas presentes en las axilas
de la hojas en cada nudo se encuentran
"dormidas", es decir en estado latente, sin
crecimiento activo. Sin embargo estas yemas
pueden reiniciar rapidamente su crecimiento si
la yema apical de eje es destruída o dañada.
Este fenómeno se conoce como "ruptura de la
dominacia apical". El mecanismo por el cual la
yema apical de un tallo controla a las restantes
yemas del eje se conoce como dominancia
apical. Estos fenómenos tienen una enorme
importancia en muchas áreas de la botánica y
las ciencias afines aplicadas como la
horticultura y jardinería.
El control que ejerce la yema apical sobre las
yemas laterales está gobernado, al menos en
parte, por la producción de Ácido Indolacético,
una hormona vegetal.
Existen casos en los cuales la yema apical
ejerce un control estrecho sobre las restantes
yemas del eje, impidiendo su crecimiento. Esto
da como resultado un patrón de crecimiento
monopodial. En este tipo de crecimiento las
yemas laterales sólo suelen activarse cuando la
yema apical está a considerable distancia.
Ejemplos típicos de este patrón son la mayoría
de las coníferas.
En cambio en otros casos la yema principal
tiene una acción más leve y no suprime el
crecimiento de las yemas laterales mas que por
un período breve. También puede suceder que
la yema principal dé origen a una flor,
perdiendo su poder de controlar a las restantes.
Este patrón de crecimiento, en el cual muchos
ejes tienen una fuerza similar se denomina
simpodial.
Por otro lado en algunas plantas crecen
predominantemente en la porción superior del
eje principal, forma de crecimiento que se
conoce como crecimiento acrótono. Este e
característico de los árboles. En cambio en
arbustos y hierbas suelen desarrollarse
activamente y hasta predominar ejes basales, lo
que se describe como crecimiento basítono.
Un tallo de crecimiento indefinido con
entrenudos desarrollados se denomina
macroblasto. Existen tallos cuyo crecimiento
es definido, sus entrenudos son muy cortos y en
su extremo se desarrollan hojas (Figura 3.3.3) o
una flor que se denominan braquiblastos.
El crecimiento inicial del tallo está generado
por la yema apical. Este tipo de crecimiento es
denominado también terminal. Sin embargo,

45 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
en cierto tipos de tallos, quedan porciones de
tejido meristemático, que es capaz de seguir
creciendo en etapas más tardías. Estos
meristemas se denominan meristemas
intercalares, y dan lugar a crecimiento
intercalar. Un ejemplo típico de crecimiento
intercalar lo representan las cañas. Tanto el
crecimiento terminal como el intercalar generan
crecimiento en longitud del tallo. Muchos tallos
tienen también crecimiento en grosor, como los
árboles. Este crecimiento lateral está generado
por meristemas laterales. Pero no todos los
tallos engrosados tienen este tipo de
crecimiento. En las palmeras, helechos
arborescentes (Figura 3.2.10.) y cicadáceas
(Figura 3.3.7.) el crecimiento definitivo en
grosor del tallo está dado por un tipo especial
de crecimiento apical en grosor generado por un
meristema primario.
Tipos de tallos y modificaciones adaptativas
Tronco: es el término aplicado al tallo
engrosado y lignificado de los árboles.
Estipites: son los tallos de las palmeras,
engrosados pero sin crecimiento
secundario. Los helechos arborescentes
(pteridofitas) y las cicadáceas
(gimnospermas) tienen tallos sin
crecimiento secundario similares a las
palmeras.
Cañas: son los tallos leñosos con crecimiento
intercalar de las gramineas.
Tallos volubles: son los tallos de crecimiento
helicoidal de ciertas plantas trepadoras
(Ipomea batatas).
Estolones: son tallos de crecimiento
plagiotrópico, paralelo a la superficie
del suelo, que se presenta en muchas
plantas herbáceas. En muchos casos
estos estolones son capaces de generar
raíces. Al penetrar el suelo éstas dan
cierta independencia a los tallos
inmediatamente asociados. De este
modo, los estolones pueden actuar
como una forma de propagación
vegetativa. [Frutilla (Fragaria
chiloensis), malezas (Cynodon
dactylon)].
Rizomas: son tallos de crecimiento
plagiotrópico pero subterráneos (pasto
ruso, sunchillo).
Tubérculos: son tallos almacenadores
subterráneos, usualmente resultantes
del engrosamiento de rizomas (papa).
Bulbos: son tallos cortos cubiertos de hojas.
Son característicos de algunas familias
(liliáces, amarilidáceas e iridáceas).
Existen distintos tipos de bulbos:
macizos (gladiolo), tunicados (cebolla),
escamosos (ajo).
Zarcillos: son tallos modificados de ciertas
plantas trepadoras (vid, Vitis vinifera,
Cissus sp.), que les permite afirmarse a
distintos tipos de soporte.
Cladodios: son tallos suculentos, fotosintéticos
de crecimiento indefinido que se
presentan con frecuencia en especies de
ambientes áridos (cactáceas). En ciertos
casos los tallos son aplanados con
crecimiento definido, denominándose
filocladios.
Pseudobulbos: En numerosas especies de
orquídeas epífitas los tallos están
engrosados, son aéreos y
fotosintéticos, como si fueran
cladodios, pero en su extremo tienen
una o más hojas.
Espinas caulinares: Algunos tallos
modificados se transforman en espinas.
Estas son espinas de origen caulinar.
Existen también espinas de origen
foliar.
Plantas acaules:. En ciertos grupos de plantas
que crecen especialmente en pastizales,
los entrenudos no se alargan nunca. El
tallo permanece siempre en un estado
extremadamente acortado. Estas
plantas se denominan acaules. En
muchos casos estas plantas acaules son
capaces de producir un entrenudo
alargado con una flor o inflorescencia
en el extremo. Este tallo floral con un
entrenudo único recibe el nombre de
escapo. Un ejemplo típico de planta
acaule es el "diente de león"
(Taraxacum officinale).

46 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Anatomía
Estructura primaria
Existe bastante variación en la estructura
primaria de los tallos, pero pueden reconocerse
tres tipos básicos de organización:
1. En algunas coníferas y dicotiledóneas, las
células procambiales elongadas y los tejidos
vasculares que derivan de ellas aparecen como
un cilindro hueco. La región exterior de ese
cilindro se denomina córtex, y la región interior
se denomina médula.
2. En otras coníferas y dicotiledóneas, los
tejidos vasculares primarios se desarrollan
como haces discretos separados dentro del
tejido fundamental. Tanto el córtex como la
médula aparecen comunicados por parénquima
interfascicular.
Figura 4.4.3. Corte transversal de tallo de
dicotilendónea con crecimiento primario de alfalfa
(Medicago sativa). Los haces vasculares se
encuentran formando un círculo en la periferia del
tallo.
3. En el tallo de la mayoría de las
monocotiledóneas y de algunas dicotiledóneas
herbáceas la organización de haces vasculares y
parénquima interfascicular es más compleja. En
vez de aparecer como un sólo anillo de haces
vasculares, éstos suelen aparecer como varios
anillos o como un sistema de haces dispersos en
el tejido fundamental.
Figura 4.4.4. Corte transversal de tallo de
monocotiledónea con crecimiento primario
(Asparragus sp.). Los haces vasculares se encutran
distribuidos más o menos uniformemente en toda la
sección del tallo.
Distintos tipos de Estela
Los tejidos vasculares primarios, xilema y
floema, y la médula cuando está presente,
forman el cilindro central o estela.
Considerando en especial la posición de xilema
y floema se reconocen distintos tipos de estela:
Figura 4.4.2. Esquemas de los diferentes tipos de
estela.

47 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Protostela: es la forma más sencilla y
probablemente más primitiva. Un cilindro
central sólido de xilema de sección circular está
rodeado de floema. No hay médula. En la
polistela hay varios de estos cilindros de xilema
rodeados de floema. Estas son estructuras
presentes en pteridófitos. Una variante más
evolucionada sería la plectostela, presente en
licopodiáceas.
Actinostela: Una variante de la protostela, con
disposición radial de xilema y floema, presente
en pteridófitos primitivos.
Sifonostela: Los tejidos vasculares se presentan
con un espacio medular o con células
parenquimáticas en el centro.
Dictiostela: Es una variante de la sifonostela,
en la cual la estructura original de ésta se
encuentra alterada por la presencia de trazas
foliares.
Eustela: Aquí la sifonostela original se halla
modificada por la presencia de lagunas foliares
y parénquima interfascicular. En la atactostela
los haces vasculares de xilema interno y floema
externo se encuentrans distribuídos más o
menos homogeneamente en la sección.
Estructura secundaria
En los helechos, la mayoría de las
monocotiledóneas y muchas dicotiledóneas
hebáceas, el crecimiento en grosor de tallo y
raíz cesan cuando se produce la maduración de
los tejidos primarios. En cambio,en numerosas
dicotiledóneas leñosas y todas las
gimnospermas el crecimiento continúa
indefinidamente aún en órganos que no se estan
elongando,.Este incremento en grosor,
denominado crecimiento secundario, es el
resultante de la actividad de dos meristemas
laterales: el cámbium y el felógeno.
En el cámbium,a diferencia de los meristemas
apicales, las células están altamente
vacuolizadas. Existen dos tipos de células, las
iniciales fusiformes, que dan origen a las
células del sistema axial, y las iniciales
radiales, que dan origen a los radios
medulares. El cámbium vascular genera hacia
afuera, floema, y hacia adentro, xilema. El
cámbium va quedando en posición más
periférica a medida que va agregando capas de
tejido xilemático y la parte central del tallo
crece,. Para adaptarse a este cambio, el anillo de
cámbium debe ir incrementando su
circunferencia.
Figura 4.4.5. Tallo de dicotiledónea con crecimiento
secundario con xilema resultante de tres años de
crecimiento (Tilia sp.).
Figura 4.4.6. Corte transversal de tallo de una
gimnosperma (Pinus sp.) mostrando los anillos de
crecimiento.
En las regiones templadas y
subtropicales el cambium vascular está en
estado de dormición durante el invierno y se
reactiva durante la primavera. En esta
reactivación las células cambiales absorben
agua, se expanden radialmente y se dividen
periclinalmente. Durante esta expansión las
paredes celulares se vuelven más delgadas.
Como resultado de este proceso, todos los
tejidos hacia afuera del cambium pueden ser
separados más facilmente del tallo. Esta
reactivación del cambium vascular está

48 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
gobernada a su vez por la reactivación de las
yemas caulinares, que comienzan a producir
hormonas de crecimiento (auxinas) que se
desplazan basípetamente, estimulando la
actividad cambial.
Durante el período de máxima
actividad del cambium, máxima fotosíntesis y
consecuentemente gran transpiración del
follaje, se producen células xilemáticas de
grandes dimensiones, capaces de transportar
volúmenes de agua suficientes para sustener la
demanda transpiratoria.
Paulatinamente, durante las etapas
finales de la estación de crecimiento, la
actividad fotosintética y transpiratoria se reduce
y el cambium comienza a generar células más
pequeñas y con paredes celulares más gruesas.
Este cambio en la estructura de las células se
manifiesta a nivel micro y macroscópico por la
formación de los llamados anillos de
crecimiento (Figuras 4.4.5. y 4.4.6.). Estos
pueden ser usados para determinar la edad de
los troncos y también para estimar las
condiciones de crecimiento durante el año en
que el anillo en cuestión se generó. Esto ha
permitido estimar las condiciones de
temperatura y/o humedad en épocas pasadas de
la historia, en momentos en los cuales no
existían registros meteorológicos. De esta
manera en Europa se han realizado
reconstrucciones climáticas que llegan a 8.000
años AP (Antes del Presente). Estas llegan a
3500 años atrás en la Patagonia. En el NOA, se
está avanzando en la reconstrucción climática
de los últimos siglos, unos 400-600 AP.
Las disciplinas que se dedican al
estudio de los anillos de crecimiento de los
árboles se denominan dendrocronología y
dendroecología.
Al aumentar la edad del árbol, la zona
central del xilema comienza a perder
funcionalidad y cesa el transporte de agua. En
muchas dicotiledóneas al producirse esta
situación las células de parénquima xilemático
empiezan a crecen por las puntuaciones y
penetran en el lumen de las células de los vasos
ocupándololos por completo. En una segunda
etapa estas células cambian su metabolismo y
comienzan a producir polifenoles y taninos, con
actividad antifúngica limitando el riesgo de
putrefacción, y provocando en última instancia
la muerte de las propias células. Este fenómeno
se denomina tilosis. Frecuentemente, este
proceso determina un marcado cambio de color
en el tejido xilemático central. El tallo queda
entonces con una región central oscura de
tejidos que ya no son conductores, más
resistente mecanicamente y a la pudrición, que
se denomina duramen. Los tejidos xilemáticos
periféricos más claros, reciben del nombre de
albura y son los que realizan el transporte de
agua y sales. (Fig. 4.4.7).
Figura 4.4.7. Tronco de Handroanthus
impetiginosus (=Tabebuia impetiginosa; lapacho
rosado, Bignoniaceae) cortado transversalmente.
Luego del corte los tejidos se han deshidratado y
contraído diferencialmente. Por las tensiones
generadas se ha producido la separación de las capas
más externas. Las zonas de rotura corresponden al
felógeno y al cambium, cuyas células se desgarran
facilmente por tener las paredes celulares muy
delgadas, carácterística típica de las células
meristemáticas.

49 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
4.5. RAIZ Concepto, funciones
En la mayoría de las plantas vasculares la raíz
constituye la porción subterránea del cuerpo
vegetativo, cuyas principales funciones son
anclaje al suelo y absorción de agua y
minerales. Otras dos funciones están
usualmente asociadas a las raíces: almacenaje
de sustancias de reserva, y conducción de agua,
minerales y carbohidratos. La mayoría de las
raíces son importantes órganos de
almacenamiento, y algunas, como por ejemplo
la zanahoria, remolacha y batata están
especialmente desarrolladas para el almacenaje
de sustancias de reserva. Sustancias
alimenticias sintetizadas en las partes aéreas, en
las porciones fotosintetizadoras de la planta se
mueven a través del tejido de conducción
(floema) hacia la raíz. Cierta proporción de
estas sustancias alimenticias son consumidas
por la propia raíz para su mantenimiento y
crecimiento, pero una proporción importante
suele ser almacenada para ser "digerida"
parcialmente después y transportada
nuevamente a las partes aéreas. En las plantas
bianuales (plantas que completan su ciclo de
vida en dos años), como la zanahoria y la
remolacha, grandes cantidades de reservas se
acumulan en la raíz durante el primer año y son
empleadas durante el segundo para producir
flores, frutos y semillas. Agua y minerales
(iones inorgánicos), absorbidos por la raíz se
mueven por el xilema hacia las partes aéreas.
Además, las raíces son fuentes muy importantes
de hormonas vegetales, en particular hormonas
que estimulan la división celular (citocininas) y
el alargamiento celular (giberelinas), o que
reducen el crecimiento y alargamiento celular, y
la transpiración (ácido abscísico). Estas
sustancias cumplen un rol esencial en la
comunicación de mensajes entre la raíz y las
partes aéreas, que dependen de estas hormonas
para su crecimiento, desarrollo y prevención de
estrés hídrico.
Origen: raíces normales y adventicias
Los sistemas radicales se clasifican
generalmente en dos tipos básicos. En el primer
tipo todo el sistema es derivado del crecimiento
y ramificación lateral de la raíz embrionaria y
se denomina sistema primario, normal, o
embrional. Este tipo se encuentra típicamente
en dicotiledóneas y gimnospermas. En un
segundo tipo, el sistema primario es suplantado
por un sistema adventicio. Este tipo es
característico de las monocotiledóneas, pero
también puede presentarse en dicotiledóneas.
Las pteridofitas también poseen un sistema
radical exclusivamente adventicio. Un sistema
adventicio se desarrolla a partir de primordios
radicales que se desarrollan sobre un tallo o
más raramente una hoja. “Adventicio” es un
término que significa “llegado desde el
exterior” y en morfología se aplica usualmente
a cualquier órgano que se encuentra en una
posición atípica. Sin embargo, esta es la
situación típica en grandes grupos como las
monocotiledóneas, en los cuales la situación
normal es que la raíz embrionaria muera y sea
reemplazada por raíces que se originan en el
tallo. Aunque el término “adventicio” no parece
lo más apropiado en ese caso, el uso lo ha
impuesto.
Raíces pivotantes y raíces fasciculadas
La primera raíz de una planta se origina en el
embrión, y es usualmente denominada raíz
primaria. En muchas gimnospermas y
dicotilendóneas la raíz primaria suele
transformarse en una raíz típica, pivotante o
axonomorfa y suele crecer directamente hacia
abajo, dando lugar a ramificaciones o raíces
laterales. Este tipo de sistema se denomina
alorrizo. Las raíces adventicias en cambio suelen
dar lugar a un sistema de raíces fibrosas o
fasciculadas, en el cual ninguna es más
prominente que las otras. Este sistema radicular se
describe como homorrizo.Los sistemas radicales
fibrosos suelen formar un entrelazado muy estable

50 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
en las capas superficiales del suelo, dando
generalmente una buena protección contra la
erosión superficial. Los sistemas alorrizos por su
parte, pueden contribuir a la estabilidad del suelo
en laderas.
Regiones de la raíz
En forma macroscópica pueden distinguirse una
serie de partes en una raíz: se denomina cuello de
la raíz a la porción por la cual esta se une al tallo,
y que usualmente se encuentra cerca del nivel del
suelo. El eje principal de la raíz suele
denominarse cuerpo y las raíces laterales se
denominan ramificaciones.
Zonas de la raíz En el extremo o ápice de la raíz también pueden
distinguirse una serie de zonas (figura 1). A veces
éstas son visibles a simple vista, pero con
frecuencia es necesario recurrir a una lupa o
microscopio. El extremo de la raíz está cubierto
por cofia o caliptra, una masa de células que
protege el punto de crecimiento o meristema y
favorece la penetración. A medida que la raíz se
alarga y empuja a través del suelo las células de la
cofia mueren y se desprenden, formando una
masa lubricante de aspecto gelatinoso. Las células
perdidas por la cofia son continuamente
reemplazadas por nuevas células generadas por el
meristema. La sustancia que forma esta gelatina
es rica en polisacáridos hidratados, en su mayor
parte de naturaleza pectínica, que es secretada por
las células de la cofia. Además de proteger el
meristema apical, la cofia cumple un importante
rol controlando la respuesta a la fuerza de
gravedad (gravitropismo). Cuando se elimina la
cofia sin dañar el meristema, usando técnicas de
microcirugía, el ápice radical pierde la capacidad
de responder al estímulo gravitrópico. El
mecanismo sensor de la gravedad parece estar
constituido por granos de almidón, llamados
estatolitos, cuya migración y deposición en un
lado particular de la célula determina la respuesta
gravitrópica.Luego del meristema, que ocupa
una porción bastante pequeña, existe una zona
donde se producen abundantes divisiones
celulares, seguida de una zona de
alargamiento, que usualmente tiene algunos
milímetros de longitud. Por encima de esta
región la raíz ya no se alarga. Por lo tanto el
crecimiento en longitud está restringido a una
región pequeña cerca del ápice.
El crecimiento de las raíces en el suelo
es un proceso relativamente continuo que sólo
se frena cuando las condiciones son adversas:
suelo muy seco, muy frío o muy compacto. El
crecimiento permite ir explorando
constantemente nuevas zonas y aprovechando
en consecuencia nuevas fuentes de nutrientes y
agua. Durante su crecimiento las raíces suelen
buscar los puntos de menor resistencia, y con
frecuencia siguen los canales dejados por raíces
muertas y descompuestas con anterioridad.
Figura 4.5.1. Zonas de la raíz: (1) zona
meristemática cubierta por la cofia, (2) zona de
alargamiento, (3) zona de absorción y maduración,

51 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
(4) zona suberificada. Modificado de Raven et al.
1992.
Por encima de la zona de alargamiento existe
una zona de maduración, donde los tejidos
internos de la raíz maduran. En esta zona
también se generan los pelos radicales, por ello
también suele ser llamada también zona de
absorción. Los pelos radicales tienen en
general una vida limitada a unos pocos días,
siendo reemplazados por nuevos pelos que se
van generando en la porción más joven. Por
encima de la zona cubierta por pelos se presenta
una porción suberificada, cuya capacidad de
absorción suele ser mucho más restringida. En
esta región, donde el crecimiento en longitud ya
ha concluido, pueden presentarse las
ramificaciones laterales. En la mayoría de las
plantas con semillas las raíces laterales se
originan endógenamente (es decir en zonas
internas), a partir del periciclo (una capa de
tejido meristemático que rodea al cilindro
central).
Anatomía de la raiz
Un corte longitudinal del ápice de la raíz
permite apreciar una serie de zonas definidas.
En el extremo distal se encuentra la cofia,
recubriendo al meristema apical. Este se halla
formado por un grupo de células
(angiospermas, gimnospermas), o por una sola
célula tetraédrica (helechos). En sentido
proximal encontramos grupos de células
derivadas del meristema, que continúan
dividiéndose activamente, en sectores
claramente definidos que dan origen a las
distintas zonas: epidermis, cortex (=corteza) y
cilindro central.
Un corte transversal de la raíz (Figura 4.5.2.)
revela dos zonas claramente diferenciadas: una
zona interna llamado cilindro central, donde se
encuentran los tejidos de conducción rodeados
por la endodermis; el cilindro central está
delimitado por el periciclo, uni a
pluriestratificado (Gimnospermas y algunas
Angiospermas, entre ellas algunas gramíneas).
Se encuentra rodeado por una zona externa,
formada por un tejido parenquimático llamado
cortex, limitado por la rizodemis con pelos
absorbentes en las porciones jóvenes de la raíz.
En las zonas más viejas la epidermis se
suberifica, o puede llegar a desaparecer al ser
reemplazada por una capa de células
suberificadas más profundas llamada
exodermis. La epidermis suberificada o la
exodermis son mucho menos permeables al
agua que las células rizodérmicas con pelos
absorbentes. En papa se ha observado que las
porciones de raíz joven con pelos absorbentes
tiene una capacidad de absorción de nutrientes
4 veces mayor que las porciones viejas.
En el cilindro central los tejidos
conductores, xilema y floema se hallan
intercalados, formado bandas. El crecimiento
secundario de la raíz se produce en
dicotiledóneas y gimnospermas por la
diferenciación de una banda de cambium entre
las bandas de floema y xilema. El cambium
genera, en forma análoga a lo que sucede en el
tallo, xilema hacia el interior y floema hacia el
exterior. Con el tiempo, el resultado de este
proceso es una estructura secundaria con anillos
anuales esencialmente idéntica a la que se
observa en los tallos con crecimiento
secundario.
Endodermis, apoplasma, simplasma y
absorción de agua
El agua y las sales minerales pueden atravesar
las paredes y membranas celulares de los pelos
absorbentes, penetrando al citoplasma. A
través de los plasmodesmas pueden pasar a
otras células y así distribuirse en toda la
planta. Por otro lado, el agua y las sales
pueden mantenerse en las paredes celulares,
sin atravesar membranas y sin entrar en
contacto directo con el citoplasma. Estas dos
posibilidades implican la existencia de dos
“espacios” dentro de una planta. Uno de ellos
“controlado” por membranas, es decir al cual
penetran sustancias que sufren en cierta
medida una selección por parte de membranas
semipermeables, y otro al cual se accede sin
atravesar ninguna membrana. El espacio
interno es la porción usualmente más rica en
nutrientes y materia orgánica, está controlado
por membranas y recibe el nombre de

52 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
simplasma. Por su parte el espacio
conformado por las paredes y espacios
intercelulares se denomina apoplasma.
La presencia de la endodermis con su banda
de Caspari implica sin embargo, que para
poder penetrar al cilindro central tanto agua
como sales se vean obligadas a atravesar la
membrana plasmática de estas células. La
deposición de endodermina, impermeable al
agua y solutos determina que la circulación
apoplástica a través de las paredes se vea
interrumpida en esa zona. La endodermina es
una sustancia lipídica químicamente afín a la
suberina y cutina.
La existencia de un límite continuo al flujo
apoplástico determina también que la raíz
pueda funcionar en cierto modo como un
osmómetro. Si la concentración de solutos es
mayor en el cilindro central que en el exterior,
situación que se da usualmente, se produce un
flujo de agua desde el exterior, es decir desde
el suelo, hacia el interior del cilindro central y
consecuentemente desde abajo hacia arriba,
hacia el follaje. Esta presión positiva que
bombea agua hacia arriba se conoce como
presión radical. Es uno de los factores que
permiten el ascenso de la savia no elaborada
desde las raíces hacia el follaje.
Dimensiones de los sistemas radiculares
La extensión del sistema radical, es decir
cuánto penetra en profundidad y cuánto se
extiende lateralmente, depende de diversos
factores, como humedad, temperatura y
composición del suelo. La mayor parte de las
raíces "activas", es decir aquellas que participan
más activamente en la absorción de agua y
minerales se encuentran en el primer metro de
profundidad. Las raíces activas de la mayoría
de los árboles se encuentran en los 15 cm
superficiales. Algunos árboles, por ejemplo los
sauces, suelen producir raíces pivotantes poco
profundas. Otros en cambio, como los pinos,
suelen producirlas muy profundas, lo cual hace
muy difícil transplantarlos pasada cierta edad.
El récord de penetración en profundidad de
raíces ha sido citado para el mesquite (Prosopis
juliflora), cuyas raíces se encontraron a 53 m de
Figura 4.5.2. Corte transversal de una raíz mostrando las vías de entrada de agua desde el suelo hacia el
cilindro central: (1) vía apoplástica atravesando paredes celulares hasta llegar a la endodermis, (2) vía
simplástica, (3) vía tanto simplástica como apoplástica.

53 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
profundidad en una mina en Arizona (Estados
Unidos). Parientes cercanos del mesquite son
los algarrobos (Prosopis alba,P. nigra), cuyas
raíces se han encontrado hasta 30 m de
profundidad en perforaciones para pozos de
agua en los Valles Calchaquíes. En las zonas
áridas las raíces profundas cumplen un rol
esencial en la estrategia de adaptación al
ambiente de la mayoría de los árboles y
arbustos. El aprovechamiento del agua en
horizontes profundos del suelo también es muy
importante en algunas regiones tropicales
húmedas. En una extensión enorme de
Amazonia (alrededor de 2 millones de km2), las
raíces presentes entre 2 y 8 m de profundidad
son fundamentales para mantener la selva con
todo su follaje transpirante durante 3-5 meses al
año, cuando las lluvias no alcanzan a cubrir la
demanda de agua.
Las raíces de la alfalfa (Medicago
sativa) pueden extenderse hasta más de 6 m de
profundidad. La extensión lateral de las raíces
suele superar la proyección de la copa. Las
raíces del maíz (Zea mays) pueden alcanzar 1.5
m de profundidad y 1 m de radio alrededor de
la planta.
Un aspecto muy importante de las
raíces es su superficie de absorción. Uno de los
estudios más detallados en este sentido
realizado en una planta centeno (Secale
cereale), ha mostrado que la superficie total del
sistema radical de una planta era de 639 m2,
aproximadamente 130 veces el área foliar, y
ocupaban un volumen total de sólo 6 litros de
suelo. La mayor parte de la superficie de
absorción de un sistema radical está dada por
los pelos radicales, que en la planta de centeno
de nuestro ejemplo totalizaron unos 14 x 109
(catorce mil millones) con una superficie de
400 m2. Si todos estos pelos se alinearan uno
detrás de otro superarían los 10.000 km. Esta
zona de raíces nuevas y en crecimiento es la
que posee mayor capacidad de absorber agua y
minerales. Por esa razón cuando se transplanta
una planta a otro lugar es conveniente hacerlo
con la mayor cantidad posible de suelo. Si la
planta es simplemente arrancada de su lugar, la
mayoría de sus raíces activas quedan allí y la
planta tiene menos probabilidades de
sobrevivir.
El sistema radical y las partes aéreas tienen
dimensiones tales que para una especie y
condiciones ambientales determinadas siguen
una proporción mas o menos fija. Por ejemplo
en el trébol blanco (Trifolium repens) la
proporción tallo/raíz es aproximadamente 2/1,
mientras que en el trebol del cáucaso (T.
ambiguum) es de 1/3. Por ello el trébol del
Cáucaso puede vivir mejor en ambientes secos.
Cuando esta proporción es alterada por causas
externas, la planta tiende a restablecerla. Si el
ganado consume el follaje, la planta da
prioridad a la regeneración de éste en
detrimento de más raíces, hasta que la
proporción original se restablece. Como ya se
dijo, cuando se transplanta una planta, una parte
sustancial de las raíces es destruida. En su
nuevo lugar, la planta suele invertir un
considerable tiempo y energía regenerando el
sistema de raíces antes de reiniciar el
crecimiento del follaje.
La capa de suelo que rodea a la raíz y se halla
íntimamente adherida a ella, conteniendo
células muertas de la cofia y abundantes
microorganismos es denominada rizósfera. Así
como la atmósfera es una capa que recubre toda
la tierra, la rizósfera es una capa que cubre la
mayor parte de la superficie de los continentes.
Por el hecho de crecer en el suelo las raíces en
general son más difíciles de estudiar que los
otros órganos. Eliminar el suelo implica con
frecuencia dañar la raíz irreversiblemente o al
menos alterarla de modo marcado. Eliminar el
suelo para estudiar una raíz es proceso
engorroso en el caso de plantas pequeñas y
sencillamente imposible en el caso de árboles
de ciertas dimensiones. Por estas razones las
raíces han sido menos estudiadas y son menos
conocidas que los otros órganos vegetales. Con
frecuencia es difícil herborizarlas
correctamente. Esto, sumando a las dificultades
para extraerlas ha determinado que las raíces
estén pobremente representadas en los
herbarios.

54 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Modificaciones adaptativas de la raíz
La mayoría de las raíces cumplen función de
almacenamiento de sustancias alimenticias en
alguna etapa de la vida de una planta. Sin
embargo en algunos casos, están especializadas
para esta función. Estas raíces suelen ser de
consistencia carnosa debido a que poseen
abundante parénquima almacenador. Estas
raíces se denominan reservantes o tuberosas.
Los ejemplos más comunes han sido
nombrados anteriormente: batata (Ipomea
batatas), zanahoria (Daucus carota), remolacha
azucarera (Beta vulgaris; en este caso el tallo
también suele formar una porción del órgano
almacenador). El yacón (Smallanthus
sonchifolius) y la ahipa (Pachyrhizus ahipa)
son dos especies originarias de la región andina
con raíces almacenadoras comestibles que
cumplen la curiosa función de “frutos” en la
dieta.
Comúnmente las raíces son
subterráneas, sin embargo existen muchos casos
de raíces aéreas o epígeas. Con frecuencia
éstas poseen cloroplastos y son capaces de
realizar fotosíntesis, recibiendo el nombre de
raíces fotosintéticas. Las plantas epífitas de las
familias de Orquídeas y Bromeliáceas suelen
poseer raíces aéreas.
Dentro de las plantas vasculares existe un
número importante de especies que viven como
parásitas de otras. Estas especies parásitas
poseen raíces especializadas que penetran en
los tejidos de otras para extraer agua y
nutrientes. Estas raíces se denominan
haustorios. Como ejemplos de plantas que
poseen haustorios pueden mencionarse a la
cuscuta (Cuscuta sp.) y la liga (Ligaria
cuneifolia).
En áreas inundadas o pantanosas la
concentración de oxígeno en el suelo suele ser
extremadamente baja, dificultando la
respiración de las raíces. Algunas especies
superan este problema produciendo raíces
modificadas que crecen en sentido vertical
(gravitropismo negativo) hasta la superficie del
agua. Estas raíces se denominan
pneumatóforos. Existen varias especies de
arboles denominados mangles (ej.: Avicennia
germinans), típicas de áreas costeras
(manglares) en las regiones tropicales que se
caracterizan por poseer este tipo de raíces.
Algunas raíces tienen la capacidad de generar
yemas apicales y dar lugar al crecimiento de
tallos, se habla en este caso de raíces
gemíferas. La raíz almacenadora de la batata
tiene aptitud gemífera. Algunos árboles, por
ejemplo algunos álamos (Populus sp.) o el
arrayán (Eugenia uniflora) poseen raíces
laterales gemíferas que cumplen un activo rol
en la propagación vegetativa de la especie.
Algunas especies con raíces de este tipo pueden
transformarse en malezas difíciles de erradicar.
En ciertos casos como en el lapacho
(Handroanthus impetiginosus =Tabebuia
avellanedae), la aptitud gemífera se desarrolla
una vez que la raíz ha sido dañada
mecánicamente.
En ciertas especies sin crecimiento en grosor
del tallo, en especial en bosques y selvas
lluviosas, suelen producirse raíces adventicias a
distintos niveles del tallo que suelen reforzar la
resistencia mecánica del mismo. Estas raíces se
denominan fúlcreas. Son comunes en especies
de helechos arborescentes y en algunas especies
palmeras y en Pandanus sp. Otras raíces de
sostén que actúan en forma similar se
encuentran también en maíz y en muchas
especies de gomero (Ficus sp.). Estas últimas
reciben el nombre de raíces columnares.
También cumplen una función de sostén las
raíces tabulares, que forman una especie de
contrafuerte en la base de tallo de algunas
especies de árboles tropicales.
En numerosas plantas bulbosas o rizomatosas
existen raíces contráctiles. Estas cumplen la
función enterrar paulatinamente a los bulbos o
rizomas, evitando que queden en superficie. La
contracción se produce por un crecimiento en
grosor que provoca un acortamiento.
Raíces simbióticas
Nódulos radiculares.
Como se mencionó más arriba, una de las
funciones principales de las raíces es absorber

55 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
agua y minerales nutritivos disueltos en ella.
Uno de los minerales más importantes en la
nutrición vegetal es el nitrógeno (N), que es un
componente esencial en la constitución de las
proteínas. Con frecuencia el N es un elemento
escaso en los suelos y su escasez relativa limita
el crecimiento vegetal. En cambio, es muy
abundante en la atmósfera terrestre. Las plantas
vasculares no son capaces de utilizar el N del
aire, mientras que distintas bacterias y
cianobacterias si pueden hacerlo. Este proceso
llamado fijación de nitrógeno, es un proceso
que requiere mucha energía química. Algunas
especies de bacterias fijadoras de N han
desarrollado una asociación con especies
vegetales, en la cual el vegetal proporciona un
medio adecuado para la vida de las bacterias,
con abundante aporte de sustancias energéticas,
y la bacteria por su parte transfiere una
proporción sustancial del N fijado a la planta
hospedante. Esta relación de beneficio mutuo
entre dos organismos diferentes se denomina
simbiosis. Las bacterias invaden porciones de
la raíz del hospedante, dando lugar al desarrollo
de nódulos radicales. Las bacterias fijadoras
más importantes en términos agronómicos
pertenecen a los géneros Rhizobium y
Bradirhizobium, que forman simbiosis con la
mayoría de las especies de la familia de las
Leguminosas, por ejemplo poroto (Phaseolus
sp.), soja (Glycine max), tipa (Tipuana tipu).
Otro grupo de bacterias filamentosas del género
Franckia, clasificadas taxonómicamente como
Actinomicetes también forman asociaciones
simbióticas y nódulos en árboles y árbustos del
género Alnus (alisos). En Tucumán existe un
representante del género (Alnus acuminata),
conocido comúnmente como aliso del cerro,
que produce este tipo de nódulos.
Si bien los primeros beneficiarios de la
simbiosis son los participantes, tarde o
temprano el nitrógeno fijado se incorpora al
suelo y puede ser aprovechado por otros
organismos. Por ello los suelos colonizados por
plantas con bacterias fijadoras asociadas suelen
incrementar su fertilidad significativamente.
Micorrizas
Más común todavía que la asociación entre
bacterias y vegetales superiores es la existente
entre hongos y vegetales superiores, que se
denomina micorriza. Los hongos carecen la
capacidad de fijar nitrógeno como las bacterias.
En cambio, son capaces de explorar volúmenes
muy importantes de suelo y solubilizar
nutrientes que de otro modo no estarían
disponibles para las plantas. Como en la
simbiosis bacteriana, la planta suministra
hidratos de carbono, mientras que el hongo
aporta minerales, en particular fósforo, y
también zinc, manganeso y cobre, todos
elementos esenciales para el crecimiento
vegetal. También se ha observado que las
micorrizas pueden contribuir a mejorar la
absorción de agua por parte de la raíz.
Existen dos tipos principales de micorrizas:
endomicorrizas y ectomicorrizas. Las
endomicorrizas se caracterizan porque el hongo
penetra las células corticales de la raíz y forma
dentro de ellas estructuras ramificadas
denominadas arbúsculos o hinchamientos
denominados vesículas. Con frecuencia este
tipo de micorriza es denominadas vesiculares-
arbusculares o VA.
Este tipo, VA, aparece en la mayoría de las
plantas herbáceas y muchas leñosas, muchos
helechos y briofitas. En este tipo de micorrizas
forman ramificaciones entre y en el interior de
las células. De esta manera se incrementa la
superficie de intercambio entre los simbiontes.
En las raíces viejas, las hifas desarrollan
vesículas terminales que acumulan lípidos
cuando maduran. Algunas leguminosas forman
una simbiosis tripartita que involucra:
micorrizas VA, nódulos bacterianos y la planta.
Por ejemplo Glycine max (soja) se asocia con
Glomus fasciculatum (hongo zigomycete) y
Bradirhyzobium japonicum (bacteria fijadora de
nitrógeno). Este tipo de asociación permite un
mejor suministro de fósforo y nitrógeno a la
planta.
Las endomicorrizas son muy comunes en los
árboles de las regiones tropicales, donde se
estima que la mayoría de las especies poseen
este tipo de asociación. Las ectomicorrizas se
encuentran particularmente en ciertos grupos de

56 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
árboles y arbustos de las regiones templadas.
Por ejemplo en la familias que incluyen a los
robles y en las de los pinos. En las
ectomicorrizas el hongo no penetra en las
células de la raíz del hospedante sino que las
rodea. Con frecuencia en las raíces con
ectomicorrizas los pelos absorbentes están
ausentes, y la función de los mismos es
desempeñada por el hongo.
Otro tipo particular de micorriza se presenta en
las Orquídeas. En esta familia de plantas el
micelio del hongo crece en parte en el interior
de las células de la raíz, en forma similar a las
endomicorrizas. El rol del hongo es
fundamental para la orquídea al punto tal que
en la naturaleza las semillas de orquídeas son
capaces de germinar y desarrollarse sólo
cuando se establece una asociación con un
hongo simbionte, que la provee de nutrientes
en las etapas iniciales de desarrollo. Recién
más tardíamente la orquídea es capaz de
fotosintetizar y volverse autosuficiente
energéticamente.
También se han descripto formas combinadas
como ecto-endomicorrizas, que son menos
comunes.
Figura 4.5.3. Visión esquemática de los distintos tipos de micorrizas en un corte transversal de raíz. Se
presentan los taxones característicos involucrados en cada tipo. V: vesícula fúngica; A: arbúsculo; Sp: espora. En
trazo grueso aparecen las estructuras del hongo. Tomado de Larcher 1995.

57 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
APENDICE: SUELOS Y NUTRIENTES MINERALES LOS HORIZONTES DEL SUELO
El suelo es una mezcla compleja que combina elementos vivos, materia orgánica en descomposición y
minerales con distinto grado de alteración. En un suelo se reconocen dos cualidades: la textura, que
está dada por el tamaño de las partículas predominantes que lo componen (de mayor a menor tamaño,
grava, arena, limo, arcilla); y la estructura, que está por la mayor o menor unión entre los distintos
materiales, formando “terrones”. En el suelo se pueden distinguir distintas capas u “horizontes”,
determinados por su cercanía a la superficie, la edad del suelo, el clima en el cual se ha formado, la
vegetación que crece sobre él y el material original en su formación. La figura 1 muestra un perfil
típico de suelo en la llanura tucumana.
Figura 1. Gentileza M. Puchulu, Cátedra de
Edafología. Facultad de Cs. Naturales e I.M.Lillo.
El Horizonte A es una capa donde se acumula
materia orgánica, por ello su color oscuro. Se
encuentra en el la mayor parte de la actividad
biológica, raíces, lombrices, bacterias, hongos,
etc.
El horizonte E es una capa de “eluviación” o
"lavado" que ha perdido materia orgánica y
arcillas, por eso es mas claro y de textura más
gruesa.
El horizonte "B" es un horizonte
iluviado o de acumulación, ya que es allí
donde se acumula lo que se lavó del "E".
El horizonte "C" es el material originario o
material parental, a partir del cual se forman
los horizontes superiores. En este caso se trata
loess, materiales finos depositados por el
viento.
Por encima del horizonte A puede presentarse un horizonte con gran cantidad de materia organica en
descomposición, hojarasca, ramas, etc., y se habla de un horizonte “O”, por organico.
LOS NUTRIENTES MINERALES Y LAS PLANTAS Elemento Forma usual de
absorción
Funciones relevantes en las plantas
Macronutrientes
Nitrogeno NO3- Constituye proteínas, bases nitrogrenadas, clorofila.
Fósforo H1PO4- Forma ácidos nucleicos, ATP
Azúfre SO42- Cofactor enzimático, forma parte de proteínas
Potasio K+ Principal electrolito celular
Calcio Ca2+ Paredes celulares, estabilizante de membranas
Magnesio Mg2+ Paredes celulares, componente de clorofila
Hierro Fe2+ Cofactor enzimático, metabolismo respiratorio celular
Micronutrientes
Zinc, cobre, molibdeno,
cloro, cobalto, níquel,
selenio, sodio,
Cofactores enzimáticos

58 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
4.6.FLOR
La flor es un eje o rama modificada para la
reproducción. Podemos definirla como un
vástago con entrenudos muy cortos, o sea un
braquiblasto, que lleva antófilos, estériles y
fértiles. Éstos últimos, los esporofilos son los
encargados de producir las esporas.
Verticilos florales
El eje de la flor, generalmente ensanchado,
recibe el nombre de receptáculo. Es la
prolongación del tallo y presenta una sucesión
de entrenudos breves en cuyos nudos se
insertan los antófilos. Éstos pueden
disponerse en forma espiralada, o sea un
antófilo por nudo (ej. Magnolia) o más
frecuentemente en ciclos o verticilos, cuando
dos o más antófilos nacen de un mismo nudo.
El número de verticilos o ciclos de las flores
cíclicas es variable. Lo más frecuente es que
existan de 4 o 5 en cada flor (flores tetra y
pentacíclicas respectivamente). De los
mismos uno corresponde, desde el interior
hacia el exterior, a las hojas carpelares, uno o
dos a los estambres y uno o dos a las hojas
periánticas.
En las flores espirocíclicas (ej.
Cactáceas), algunas piezas se disponen en
forma espiralada y otras en forma cíclica.
La flor es completa si posee todos los
verticilos (Figura 4.6.1), y es incompleta si
falta algunos de ellos.
El número de piezas florales es de 3 en
cada ciclo en las Monocotiledóneas (flores
trímeras), y generalmente 5, más raramene 4,
y hasta 6 en las Dicotiledóneas (flores
tetrámeras, pentámeras y hexámeras
respectivamente).
Figura 4.6.1. Esquema de una flor completa.
Perianto
Los verticilos estériles, cáliz y corola,
constituyen el perianto. En algunos casos no
están presentes, y se habla entonces de flores
aclamídeas o desnudas. Cuando están
presentes pueden darse dos casos:
flores monoclamídeas, cuando hay un
solo ciclo periántico representado por
el cáliz que puede ser verde o
coloreado.
flores diclamídeas, cuando poseen los
dos ciclos periánticos o sea cáliz y
corola.
En ese segundo caso se distinguen:
flores heteroclamídeas, cuando tienen
cáliz y corola de diferente color.
flores homoclamídeas, cuando no hay
diferenciación entre cáliz y corola. En
este caso se habla de perigonio.
Las piezas del perigonio se denominan
tépalos. Cuando los tépalos son de color
verdoso, es decir que tienen aspecto de cáliz,
se dice que la flor presenta perigonio calicino.
Si en cambio los tépalos se asemejan una
corola, se dice que la flor tiene perigonio
corolino.
Cáliz

59 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
El cáliz es el ciclo externo del perianto,
constituido por sépalos, es generalmente
verde y su función es la protección de los
demás ciclos florales, principalmente en el
estado de botón floral o capullo, cuando los
antófilos fértiles aún se encuentran
inmaduros. A veces es coloreado y se asemeja
a la corola. En este caso se denomina
petaloide. Los sépalos pueden estar libres
entre sí (cáliz dialisépalo) o soldados (cáliz
gamosépalo). En este último caso forman un
tubo en cuyo borde superior suelen
distinguirse los lóbulos, segmentos o dientes
que corresponden a los sépalos soldados.
Con respecto a la duración del cáliz,
este puede ser caduco, o sea que cae en la
antesis (apertura de la flor), como en la
amapola (Papaver sp.); o persistente, es decir
que permanece en el fruto y pudiendo
participar luego en la constitución del mismo
(Ej. frutilla, Fragaria sp.).
Corola
La corola es el ciclo interno del perianto
constituida por los pétalos, es generalmente
coloreada y su función principal es la de
atraer a los polinizadores. Raramente se
parece a un cáliz y en ese caso se denomina
sepaloide. Al igual que las piezas de cáliz, los
pétalos pueden estar libres entre si (corola
dialipétala) o soldados (corola gamopétala).
Corolas dialipétalas típicas son las de
las Crucíferas y las Leguminosas
papilionadas. En el primero de los casos,
como por ejemplo el rabanito (Raphanus
sativus) y el alelí (Mathiola incana), los
cuatro pétalos se disponen en forma de cruz.
En el caso de las Leguminosas papilionadas,
como el ceibo (Erythrina spp.) o la arveja
(Pisum sativum), las flores poseen 5 pétalos,
el superior y más grande se denomina
estandarte, los dos laterales alas y los dos
inferiores que se encuentran soldados
constituyen la quilla (Figura 4.6.2).
Figura 4.6.2. Flor papilionada típica de
Pisum sativum (arveja), con los pétalos
separados (izquierda) y en su posición natural
en la flor (derecha).
Las distintas formas que adoptan las corolas
gamopétalas son importantes en la taxonomía
de algunos grupos de plantas. Los tipos más
comunes son los siguientes (Figura 4.6.3):
Rotácea: tubo corto y limbo (parte
extendida) en forma de rueda o
estrella (Ej. Primula, Solanum).
Personada: corola bilabiada, cuyo
labio inferior tiene una abolladura que
cierra la garganta corolina (Ej.
Conejito, Anthirrinummajus).
Tubulosa: su forma es practicamente
cilíndrica (Ej. palán-palán,
Nicotianaglauca; flores del disco de
las Compuestas).
Campanulada: en forma de campana
(Ej. Campanula sp.).
Hipocrateriforme: tubo largo y
angosto, limbo plano y extendido (Ej.
Petunia sp.).
Infundibuliforme: en forma de
embudo (Ej. campanilla, Ipomoea sp.).
Urceolada: en forma de olla, con tubo
largo y ventricoso y el limbo poco
desarrollado (Ej. huevito de gallo,
Salpichroaoriganifolia).
Labiada: poseen dos labios, más raro
uno (Ej. Salvia sp.).
Ligulada: poseen un tubo de longitud
variable que se continúa en un limbo
mas o menos expandido en forma de
lengua (Ej. flores marginales de los

60 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
capítulos de algunas Compuestas
como la margarita, Chrysanthemum
sp. y el girasol, Helianthus annuus).
Figura 4.6.3. Tipos de corola gamopétala.
La duración de la corola es variable,
en muchos casos es breve, y por lo general se
marchita y cae después de la polinización.
La disposición de las piezas del cáliz y
la corola en el pimpollo o botón floral
constituyen la prefloración, de la cual hay
distintos tipos(Figura 4.6.4):
Valvar: las piezas de un mismo
verticilo sólo se tocan los bordes.
Contorta o torcida: todas las piezas
se cubren en parte. Cada pieza cubre a
la inmediata y es cubierta por la
precedente, Es decir tiene un lado
interno y otro externo.
Imbricada: hay una pieza interna que
no es cubierta por ninguna de las
otras, una interna que es cubierto en
ambos márgenes y las otras cubiertas
en uno de los márgenes solamente.
Figura 4.6.4. Tipos de prefloración.
VERTICILOS FÉRTILES
Androceo
Es el verticilo floral compuesto por los
estambres o microsporofilos. En
Dicotiledóneas que conservan caracteres
primitivos (Ej. Magnolia grandiflora), los
estambres tienen forma de hoja con los
microsporangios dispuestos sobre la cara
adaxial de las mismas.
Un estambre típico de Angiosperma
consta de dos partes: una fértil, la antera y
una estéril, el filamento (Fig. 5).
Generalmente una antera está constituida por
dos tecas (ditécicas), separadas entre sí por un
tejido estéril, el conectivo. En el filamento y
conectivo existe un solo haz vascular.
Figura 4.6.5. Estambres y tipos de inserción
de las anteras en el filamento.
En Malváceas, por ejemplo, las
anteras son monotécicas, es decir tienen una
sola teca.
En un corte transversal de una antera
joven (Figura 4.6.6) se observa que cada teca

61 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
está constituida por dos microsporangios
osacos polínicos que contienen las células
madres de los granos de polen. Estos se hallan
separados entre sí por un tabique constituido,
en gran parte, por tejido parenquimático.
Figura 4.6.6. Esquema de corte transversal de
una antera y de un grano de polen (escalas
diferentes).
Desde adentro hacia fuera, la pared de
los microsporangios consta de las siguientes
capas o estratos:
Tapete: rodea a las células madres de los
granos de polen, sus células poseen bastante
citoplasma y uno o varios núcleos. Tiene
como función nutrir a las microsporas en
formación. Según su comportamiento se
diferencia el tapete secretor cuando las
células mantienen su individualidad mientras
mientras viven y el tapete ameboidal,en el
que las paredes celulares se destruyen, de
modo que los citoplasmas y los núcleos
forman una masa que rodea a las microsporas.
Estratos parietales: rodeando al tapete hay
una serie de capas de células que constituyen
los estratos parietales. Estas células son
aplanadas y tienen una corta vida ya que son
aplastadas y mueren durante la maduración de
la antera.
Endotecio: se halla inmediatamente debajo
de la epidermis, se diferencia como estrato
mecánico a la madurez de la antera. Consta de
una o varias capas de células.
Exotecio: es el estrato más externo y consta
de típicas células epidérmicas cubiertas con
cutícula.
Según el número de estambres que
poseen las flores se clasifican en monandras,
diandras o poliandras si tienen uno, dos o
más estambres respectivamente.
Según el número de estambres en
relación con el número de piezas en cada
verticilo del perianto, las flores pueden ser:
Isostémonas: el número de estambres es
igual al de los pétalos.
Diplostémonas: el número de estambres es
igual al doble del número de pétalos.
Meyostémonas: el número de estambres es
menor que el número de pétalos.
Según la longitud de los estambres se
diferencian los siguientes tipos de androceos
(Figura 4.6.7):
Didínamos: cuando hay dos estambres más
largos por ejemplo en Labiadas (peperina y
menta, Mentha spp.; salvia, Salvia sp.) y
Bignoniáceas (lapacho rosado, Handroanthus
impetiginosus = Tabebuia avellanedae).
Tetradínamos: cuando son 4 estambres más
largos que los restantes, por ejemplo en
Crucíferas (berro, Rorippa nasturtium-
aquaticum; rabanito, Raphanus sativus.)
Figura 4.6.7. Tipos de androceo según la
longitud del filamento y la posición relativa
de los estambres.
Los estambres pueden estar
fusionados de diversas maneras (Figura
4.6.8):
Androceo sinantéreo: estambres unidos por
las anteras. Ej. Compuestas.
Androceo monadelfo: la unión se realiza por
los filamentos formando un solo haz. Ej.
Malváceas, rosa china, Hybiscus rosa-
sinensis).
Androceo diadelfo: los filamentos se hallan
soldados entre si formando dos grupos (9
soldados y uno libre). Ej. Leguminosas
papilionadas: arvejilla, Lathyrus odorata;
poroto, Phaseolus vulgaris.

62 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Androceo poliadelfo: los estambres se hallan
unidos por los filamentos formando varios
haces o manojos. Ej. Citrus sp.
Androceo sinfiandro: los estambres se
hallan unidos entre si por antera y filamento.
Ej. palo borracho, Ceiba insignis; zapallo,
Cucurbita maxima.
Por otra parte los estambres pueden
unirse con la corola (estambres epipétalos),
en forma total o sólo en la base (Ej. jazmín
magno, Plumeria rubra; campanilla, Ipomoea
sp.). También pueden estar soldados con el
gineceo, formando una columna llamada
ginostemo (Ej. orquídeas).
Según el lugar en que el filamento se une a la
antera, ésta puede ser (Figura 4.6.5):
Basifija: unida por la base.
Apicifija: unida por el ápice.
Dorsifija: unida por el dorso.
Versátil: la antera está sujeta al filamento
sólo por un punto y oscila alrededor de éste.
La apertura de las tecas para dejar en libertad
al polen, es la dehiscencia de la antera, que
puede ocurrir de diversas maneras (Figura
4.6.9): Dehiscencia longitudinal: se abre por
una línea lateral que sigue el eje mayor de la
teca. Dehiscencia poricida, por medio de
poros generalmente ubicados en el ápice de
las tecas. Dehiscencia valvar, por aberturas
que se producen en cada teca a modo de
valvas.
Figura 4.6.8. Distintos tipos de unión de los
estambres.
Figura 4.6.9. Distintos tipos de dehiscencia
de las anteras.
Estructura del grano de polen
Los granos de polen pueden tener
formas variables (esférica, elipsoidal, etc.) y
normalmente están protegidos por una capa
externa constituida por varias subcapas, la
exina (Figura 4.6.6), muy resistente y
formada por sustancias terpenoides
(esporopolenina) que protege el contenido
vivo del polen. En la superficie de la exina
existen diversas esculturaciones, así como
repliegues (colpos) donde la pared es más
delgada y también poros abiertos o con un
pequeño tapón u opérculo. Mas internamente,
y adosada a la exina, hay una segunda capa,

63 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
la intina, de naturaleza celulósica y péctica y
cuya función será formar el tubo polínico que
saldrá por los poros o colpos. Las diversas
formas de los granos de polen, sus distintas
esculturaciones, el número de colpos o poros,
etc., permiten identificar a los grupos a los
que pertenecen las especies que los producen.
Además las características químicas de la
exina hacen que el polen deje un claro
registro fósil que resulta muy útil en estudios
paleobotánicos, geológicos, y ecológicos. El
grano de polen maduro está formado por dos
células. Una de ellas, la mayor (Figura 4.6.6),
es la llamada célula vegetativa, y la mas
pequeña es la célula generativa, que
producirá luego los gametos masculinos.
Gineceo
Es el verticilo floral fértil mas interno
y está constituído por hojas modificadas
llamadas carpelos o megasporofilos.
Producen megasporangios llamados
óvulos,que se ubican en la cara adaxial de los
carpelos y a su vez producen en su interior
megasporas. Uno de los caracteres distintivos
de las Angiospermas es tener los óvulos
encerrados por los megasporofilos o carpelos,
debido a ello los granos de polen no germinan
directamente sobre ellos, sino que lo hacen a
cierta distancia sobre el estigma. En las
Gimnospermas los óvulos se encuentran
"desnudos" debido a que el carpelo no se
halla plegado sobre si mismo y no los encierra
por lo que los granos de polen germinan
directamente sobre ellos.
Un gineceo es unicarpelar cuando
está formado por un solo carpelo y es
pluricarpelar cuando está constituído por
más de uno.
Gineceo apocárpico o dialicarpelar
es aquel que se halla formado por varios
carpelos libres entre sí y gineceo sincárpico o
gamocarpelar es el que se encuentra formado
por varios carpelos soldados entre sí.
En las flores más evolucionadas en el
gineceo se diferencia una parte proximal
fértil, el ovario, que lleva los óvulos, una
parte distal que recibe el polen, el estigma y
una porción intermedia estéril llamada estilo a
través del cual llega el tubo polínico hasta el
óvulo (Figura 4.6.10).
El estilo puede tener distintas
posiciones con respecto al ovario (Figura
4.6.11). Es terminal cuando continúa al ovario
en su parte superior. En otros casos se
encuentra alojado en una depresión del
ovario, en este caso se trata de un estilo
ginobásico. El estigma es sésil o sentado
cuando el estilo se halla ausente.
Una flor puede ser epígina, hipógina
o perígina si las piezas del perianto y los
estambres nacen o aparentan nacer por arriba,
por debajo o alrededor del ovario
respectivamente (Figura 4.6.12).
Figura 4.6.10. Partes del gineceo.
Figura 4.6.11. Tipos de gineceo: ginobásico
(izquierda), sésil (derecha).
Muchas flores poseen hipanto que es
una estructura en forma de tubo o copa que
rodea al ovario y resulta de la prolongación
del receptáculo o por fusión de los verticilos
externos al gineceo.

64 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Figura 4.6.12. Tipos de flor según la posición
relativa del ovario y el hipanto (h).
Si el hipanto cubre por completo al ovario
soldándose a él, el ovario es ínfero y la flor se
denomina epígina. Si el hipanto está libre del
gineceo el ovario es súpero y la flor es
perígina (Figura 4.6.12). Cuando no hay
hipanto, las piezas de los restantes verticilos
nacen por debajo del gineceo, el ovario es
súpero y la flor hipógina.
Los óvulos se hallan unidos a los
carpelos en una zona especializada llamada
placenta. Las distintas formas en que ésta se
dispone se denomina placentación y puede ser
(Figura 4.6.13):
Marginal: los óvulos se disponen en doble
hilera en los márgenes de los carpelos libres.
Parietal: las placentas se hallan formadas por
las márgenes de los carpelos contiguos. En
este caso el ovario es unilocular.
Axilar: las placentas están formadas por las
dos márgenes del mismo carpelo que se ponen
en contacto en el centro del ovario. El ovario
tiene entonces tantos lóculos como carpelos.
Central: deriva de una placentación axilar en
la cual los septos o tabiques han desaparecido.
Los óvulos quedan unidos a una columna
central.
Basal: la placenta se halla restringida a la
base interna del ovario.
Apical, la placenta se encuentra en la parte
superior de la cavidad ovárica.
Figura 4.6.13. Tipos de placentación.
Estructura del óvulo o megasporangio
(Figura 4.6.14)
Muchos óvulos poseen un solo
tegumento pero otros poseen dos: la primina
y la secundinaque rodean el nucelo,que
constituye el cuerpo del óvulo, y que se
interrumpen en un punto para dar lugar a una
abertura llamada micrópila. El óvulo se halla
unido a la placenta por medio de un pie
llamado funículoel cual se halla recorrido por
un haz vascular hasta un punto llamado
chalaza. Dentro del nucelo se halla incluido
el saco embrionario constituido por 7
células: la ovocélula u oósfera (gameta
femenina) rodeada por 2 células llamadas
sinérgidas, una célula central binucleada
(losnúcleos polares) y 3 células situadas en el
polo contrario a la ovocélula, las antípodas.

65 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Figura 4.6.14. Estructura de un óvulo
anátropo.
Figura 4.6.15. Tipos de óvulos. (a) ortótropo;
(b) anátropo; (c) campilótropo.
Si el funiculo, chalaza y micrópila se hallan
orientados sobre un mismo eje se habla de un
óvulo ortótropo. Si estos se hallan formando
un ángulo de casi 180º, con la chalaza
formando el vértice se llama al óvulo
anátropo, la forma más común. Una forma
intermedia con rotación parcial es el óvulo
campilótropo.
Flor y sexualidad
Una flor es perfecta (bisexual o
hermafrodita) si posee los dos verticilos
fértiles y es imperfecta o diclina si solo
presenta uno de ellos. En este caso es
carpelada o pistilada (femenina) si no hay
estambres o si éstos son estériles, y es
estaminada (masculina) si el gineceo está
ausente o atrofiado.
Cuando las flores son imperfectas y se
disponen sobre un mismo individuo, la planta
es diclinomonoica (ej. Maíz, Zea mays). Es
diclinodioica cuando las flores imperfectas
están dispuestas sobre individuos diferentes,
uno carpelado o pistilado y otro estaminado
(Ej. sauce, Salix spp.).
Son especies polígamas aquellas que tienen
flores perfectas e imperfectas. Estas pueden
ser polígamo - dioicas si se presentan flores
carpeladas y perfectas en un individuo y
estaminadas y perfectas sobre otro diferente, y
polígamo - monoicas si tienen flores
perfectas e imperfectas en el mismo
individuo.
Dentro de las Angiospermas la mayor
proporción de especies (71%) poseen flores
bisexuales o hermafroditas, sin embargo
existen varias otras alternativas detalladas en
la tabla 4.6.1.
SIMETRÍA FLORAL Y DIAGRAMA FLORAL
Flor simétrica es aquella que puede
ser atravesada por lo menos por un plano
vertical en dos mitades simétricamente
iguales. Dentro de las simétricas hay dos tipos
principales: zigomorfa o bilateral y
actinomorfas o radiadas, según existan uno
o dos planos que dividan a la flor en dos
mitades especularmente simétricas.
Raramente existen también flores
asimétricas, que no aceptan ningún plano de
simetría (Ej. achira, Canna sp.).
El diagrama floral es una manera de
representar esquemáticamente en un plano los
diferentes ciclos que forman la flor y las
relaciones que existen entre ellos,
particularmente si se encuentran soldados o
en contacto entre sí y sus posiciones relativas.
La fórmula floral es una manera
abreviada de expresar por medio de símbolos,
letras y números la estructura de la flor.

66 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Expresión del sexo Sexo en la flor Sexo en el
individuo
Frecuencia
aproximada
Hermafroditismo ♀ y ♂ ♀ y ♂ 71 %
Monoecia ♀ ó ♂ ♀ y ♂ 5 %
Ginomonoecia ♀ y ♂/ ó ♀ ♀ y ♂ 4 %
Andromonoecia ♀ y ♂/ ó ♂ ♀ y ♂ 2 %
Dioecia ♀ ó ♂ ♀ o ♂ 5 %
Ginodioecia ♀ y ♂/ ó ♀ ♀ y ♂/ ó ♀ 7 %
Poligamia ♀ y ♂/ ó ♂/ó ♀ ♀ y ♂/ ó ♂/ó♀ 4 %
Tabla 4.6.1. Frecuencia aproximada de las diversas formas de expresión del sexo en las
angiospermas..

67 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Agente
polenizador Tipo de flor
Color de los
pétalos Polen Perfume Nectar Retribución
Coleópteros: se
piensa que fueron
los polenizadores
de las primeras
angiospermas.
grandes o en
inflorescen-
cias
grandes
blancas o de
color pálido
abundan
te
muy
perfumadas,
fragancia a
fruta, picante,
fermentación
poco o
nulo
polen, piezas
florales; el
ovario suele
estar
protegido
(ínfero)
Himenópteros:
son el grupo más
importante de
polinizadores.
con
plataformas
de aterrizaje,
con
estructuras
complejas
brillantes,
azules,
amarillas,
blancas,
reflejan UV,
con “guías de
polen”; rara
vez rojas
-- perfumes
agradables
abundan
te polen, néctar.
Dípteros --
colores
oscuros, o
blanquecinos
pálidos
-- olor
desagradable
abundan
te néctar
Lepidópteros
nocturnos
corola
alargada,
tubulosa
blanco,
amarillo
pálido
--
perfume
intenso
durante la
noche
abundan
te néctar
Lepidópteros
diurnos
corola
alargada,
tubulosa
rojo, naranja
o amarillo
brillante
-- perfume dulce
durante el día
abundan
te néctar
Pájaros
grandes o
numerosas, a
veces
colgantes
rojo, naranja,
brillante -- sin perfume
muy
abundan
te
néctar
Murciélagos
grandes, a
veces
colgantes
Blanco,
amarillo
pálido
--
perfume fuerte
a fruta o
fermentación
muy
abundan
te
néctar, polen,
piezas
florales
Viento pequeñas,
inconspicuas
Verde,
marrón, poco
vistosos
muy
abundan
te no
pegajos
o
sin perfume ausente --
Tabla 4.6.2. Características florales según el tipo de polinización.
POLINIZACIÓN
Polinización es el proceso de transporte
del polen entre los estambres y el estigma.
En las Gimnospermas este proceso se da
exclusivamente por acción del viento. Este
tipo se denomina anemofilia. Aunque en
las Angiospermas existen especies
anemófilas, la gran mayoría de ellas
tieneinteracciones, a veces notablemente
complejas con distintos tipos de animales
y se habla de zoofilia. Los insectos
(entomofilia) son los polinizadores más
importantes. Pájaros (ornitofilia) y en
menor medida murciélagos

68 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
(quiropterofilia) también cumplen ese rol
en determinados grupos de plantas.
A lo largo de millones de años
polinizadores y plantas han establecido
relaciones con frecuencia muy estrechas, y
se han influido mutuamente, resultando en
un proceso de coevolución. La tabla 4.6.2
sintetiza los distintos tipos de polinización
y las características (síndromes) que
poseen las plantas según el tipo de
polinizadores a que se encuentran
adaptadas.
INFLORESCENCIAS
Todo sistema de de ramificaciones que
remata en flores se llama inflorescencia y
el comienzo de la misma está dado por el
lugar de inserción del último nomófilo. Las
inflorescencias son casos particulares de los
distintos modos de ramificación y en ellas
suelen desarrollarse la mayoría de las
yemas ubicadas en las axilas de las hojas
tectrices, por lo que el sistema de ramas se
puede volver muy complejo y presentar
gran variedad de formas.
Desde el punto de vista evolutivo, la
existencia de inflorescencias puede
interpretarse por su atractivo mayor para
los polinizadores.
Según su sistema de ramificación
se clasifican en: racimosas, de
crecimiento indeterminado, que
usualmente culminan en una yema activa y
que en ciertas ocasiones puede reasumir el
crecimiento como yema vegetativa;
cimosas, con crecimiento determinado,
cuya yema terminal muere o se transforma
en una flor. En las inflorescencias cimosas
el crecimiento es continuado por yemas
laterales, que a su vez se transforman en
flores. En ciertos casos este tipo de
crecimiento determina una infloresecencia
con una apariencia de indeterminada, es
decir racimosa.
Para interpretar adecuadamente las
inflorescencias es muy importante tener en
cuenta:
la posición de las brácteas u hojas
tectrices.
la posición de de las yemas, flores
y frutos resultantes
la secuencia de floración, es decir
si ésta se produce desde la base
hacia el ápice de la rama en
cuestión o a la inversa.
En una inflorescencia pueden
distinguirse, además de las flores, las
siguientes regiones: el pedúnculo, que es
la parte del tallo que soporta al raquis o al
receptáculo común, y que se inicia en el
último nomófilo, terminando al aparecer
los primeros pedicelos o ramificaciones
del raquis. El eje o raquis es la parte del
tallo donde se insertan las flores. Cuando
es aplanado, como en los capítulos, se
denomina receptáculo. El pedicelo es la
ramificación que sostiene a cada flor. A
veces es muy corto o nulo y la flor resulta
sentada o sésil. Los hipsófilos o brácteas
pueden ser verdes o coloreados y son las
hojas tectrices de las yemas que
producirán las ramificaciones de las
inflorescencias o las flores mismas. En
algunos casos están modificadas para
atraer polinizadores, proteger a las flores y
frutos, etc., y entonces reciben nombres
especiales como espatas en las Aráceas
ypalmeras; glumas y glumelas en las
Gramíneas, involucros en las Compuestas,
etc.

69 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Figura 4.6.16. Inflorescencias racimosas.
Las flores están representadas
esquemáticamente por círculos. El tamaño
de los círculos indica el grado de
desarrollo. La flecha en el extremo indica
una yema terminal activa.
Inflorescencias racimosas
El racimo es la inflorescencia racimosa
básica, en la cual cada flor tiene su propio
pedicelo que nace en la axila de una hoja
tectriz y las flores maduran de abajo hacia
arriba (Ej. arvejilla, Lathyrus odorata). A
veces el raquis del racimo aparece
ramificado, constituyendo un racimo
compuesto o panícula. La espiga es
similar a un racimo sólo que las flores son
sésiles, es decir carentes de pedicelo. El
espádice puede considerarse una espiga
cuyo eje es engrosado y carnoso, que se
encuentra asociado a una bráctea
frecuentemente coloreada. Es la
inflorescencia característica de las Aráceas
(Ej. cala; Zantedeschia aethiopica). El
amento puede definirse como un racimo o
espiga péndula (Ej. sauce llorón, Salix
babylonica). La espiguilla es una
inflorescencia típica de las Gramíneas o
Poaceae, constituida por un par de brácteas
(glumas) de toda la unidad y brácteas que
protegen a cada flor, la palea y la lemma.
Usualmente la espiguilla es la unidad que
integra inflorescencias más complejas. Por
ejemplo la espiga de espiguillas o espiga
compuesta es la inflorescencia del trigo
(Triticum aestivum) y el centeno (Secale
cereale).
En el corimbo los pedicelos de las flores
basales son más alargados, de manera que
todas las flores quedan casi a una misma
altura (Ej. corona de novia, Spiraea
cantoniensis). En la umbela todos los
pedicelos son aproximadamente iguales,
pero el eje principal de la inflorescencia
está totalmente reducido, de modo que
ellos salen casi de un mismo punto.
Existen umbela de umbélulas, es decir una
umbela compuesta. Esta la inflorescencia
característica de las Umbelíferas o
Apiaceae (Ej. zanahoria, Daucus carota).
En el capítulo el eje de la inflorescencia
está engrosado, dejando una superficie
plana, convexa o cóncava rodeada de
brácteas, sobre la cual se insertan las
flores. Éstas tienen un sentido de floración
centrípeto. Es la inflorescencia
característica de la familia de las
Compuestas o Asteraceae (Ej. girasol,
Helianthus annuus).
A pesar de la diversidad, puede observarse
que todas las variantes responden al patrón
del racimo típico, ya que de éste podría
pasarse al corimbo por modificación de la
longitud de los pedicelos; del corimbo a la
umbela, por acortamiento de los entrenudos

70 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
del raquis, y de ésta al capítulo, por
reducción máxima de los pedicelos.
Inflorescencias cimosas
En estas inflorescencias la ramificación es
del tipo simpodial, de modo que el eje
principal termina su crecimiento, ya sea
porque muere o porque origina una flor, y
las ramas que de él se derivan se ramifican
también de modo simpodial. Las cimas
tienen aspecto distinto según sea el
número de yemas laterales que se
desarrollan: si solo lo hace una de ellas, se
tiene un monocasio, que cuando se
ramifica hacia un solo lado se denomina
cincinio, o cima escorpioide, mientras
que cuando la ramificación se alterna de
un lado al otro recibe el nombre de cima
helicoide (Ej. Hemerocallis sp.). Cuando
aparecen dos ramas por debajo de la flor
terminal, se la llama dicasio (Ej.
Kalanchoe sp.)y si existen varias
pleiocasio (ej. malvón, Pelargonium
hortorum). En este último caso se
denominan cimas umbeliformes pues son
similares a las umbelas pero su sentido de
floración es de adentro hacia fuera. Un
tipo particular de cima es el ciatio,
característico del género Euphorbia, que
posee flores unisexuales, frecuentemente
sin perianto. Las flores pistiladas están
reducidas a un ovario con estigma trífido,
mientras que las estaminadas son apenas
un estambre articulado al receptáculo de la
inflorescencia. Estas flores se encuentran
sobre un receptáculo cóncavo en forma de
copa y con frecuencia acompañadas de
hipsófilos vistosos.
Figura 4.6.17. Inflorescencias cimosas.
Otra inflorescencia cimosa similar es el
sícono característica del género Ficus.
Consiste en un receptáculo común,
globoso, ahuecado y suculento, que lleva
en su interior flores diclinas. Generalmente
las flores estaminadas se ubican cerca del
poro u ostíolo; las carpeladas ocupan el
resto del receptáculo y son las primeras en
abrir.
Con frecuencia algunas inflorescencias son
mixtas y puede haber en ellas partes
racimosas y otras cimosas combinadas de
diverso modo. Así puede haber cimas de
capítulos en algunas Compuestas o
racimos de cimas.

71 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
4.7. FRUTO
INTRODUCIÓN
El fruto es un órgano típico de las
Angiospermas, resultante del desarrollo del
ovario. En su forma más sencilla, el fruto es
simplemente un ovario desarrollado y maduro
luego de la fecundación (frutos genuinos) pero
muchas veces el ovario es acompañado por
otras estructurasflorales o extraflorales (frutos
complejos). Del mismo modo que las flores
han evolucionado en relación a sus agentes
polinizadores los frutos han evolucionado
adaptándose a los distintos agentes dispersores.
Al definir el fruto como un ovario
transformado luego de la fecundación se
excluye de considerar como frutos a las
estructuras que acompañan a las semillas de las
Gimnospermas. Sin embargo, muchos autores
las agrupan en seminíferas y estrobilíferas. En
el primer grupo las hojas carpelares suelen estar
poco desarrolladas, muchas veces consiste en
un simple pedicelo que soporta las semillas. En
las estrobilíferas las hojas carpelares se insertan
sobre un eje y se encuentran asociadas a
escamas tectrices.
En los frutos genuinos es muy relevante
el desarrollo de cada una de las capas de tejido
carpelar. Una hoja carpelar posee originalmente
una epidermis superior (o interior), un mesófilo
y una epidermis inferior (o exterior). Al
desarrollarse el fruto la epidermis superior da
origen al endocarpio, el mesófilo al
mesocarpio y la epidermis inferior al
exocarpio. Cada una de estas capas sufre
cambios variados que otorgan características
específicas a cada fruto yen conjunto
constituyen lo que se se denomina pericarpio,
al cual pueden agregarse, con distintos grados
de concrescencia y modificación, otras piezas
florales o extraflorales. Esta complejidad de
variantes ha dado lugar a clasificaciones de los
frutos que son en parte morfológicas y en parte
relacionadas con el mecanismo de dispersión de
las semillas. Estas clasificaciones no siempre
muestran coherencia.
CRITERIOS PARA LA CLASIFICACIÓN DE LOS
FRUTOS
Los frutos son órganos notablemente
diversos lo cual ha derivado en una gran
variedad de nombres para designarlos, muchos
de los cuales se continúan aplicando por
razones históricas. Debido a ésto, la
clasificación de los mismos puede calificarse de
artificial, es decir que no sigue necesariamente
criterios de parentesco entre especies. Especies
de familias distintas pueden tener frutos
similares y especies dentro de una familia
pueden tener frutos muy diferentes. Sin
embargo los frutos son con frecuencia un
elemento muy importante para clasificar a las
plantas.
Existen una serie de criterios para
clasificar los frutos, entre los más importantes
están:
si derivan de una sola flor (monotalámicos)
o de una inflorescencia (politalámicos).
si incluyen solamente tejidos del ovario
(genuinos) o también otras estructuras
florales o extraflorales (complejos).
si deriva de un ovario uni o gamocarpelar
(unipistilado) o dialicarpelar
(pluripistilado).
si son carnosos o secos.
si se abren a la madurez (dehiscentes) o
permanecen cerrados (indehiscentes).
Los frutos monotalámicos unipistilados,
también llamados simples, constituyen el grupo
más numeroso y variado. Entre ellos
encontramos:
FRUTOS SECOS DEHISCENTES
Legumbre: fruto monocarpelar de dehiscencia
sutural doble, común en muchas leguminosas
(poroto, arveja, soja).
Folículo: es monocarpelar de dehiscencia
sutural simple (ej. Zanthoxylon coco,
cochucho; Anadenantheracolubrina var. cebil,
cebil colorado).
Silícua: formado por dos carpelos, se parece
superficialmente a la legumbre, pero posee un
falso tabique llamado replum de origen

72 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
placentario y un marco sobre el cual se insertan
las semillas (ej. muchas Crucíferas, repollo,
alelí). Presenta dehiscencia placentífraga o
sea que el fruto se abre por medio de dos
hendiduras paralelas, muy próximas a las
placentas.
Cápsula: formado a partir de un ovario
súpero, constituido por dos o más carpelos.
Según su dehiscencia puede ser: Septicida: cuando la apertura ocurre por
separación de las paredes carpelares que
forman los septos, es decir los tabiques que
separan los lóculos. Proviene de ovarios con
placentación axilar. Loculicida: la rajadura
se produce a lo largo de la vena media de los
carpelos. Puede darse en frutos uniloculares
(Ej. Viola odorata, violeta; Viola tricolor,
pensamiento) o pluriloculares (Allium
cepa,cebolla). El fruto puede permanecer
entero o fragmentarse en valvas,
correspondiendo cada una a las mitades de dos
carpelos contiguos; cuando la placentación es
axilar, cada valva presenta el septo en su parte
media.
Septífraga: cuando la ruptura tiene lugar en
los septos o tabiques, sobre planos paralelos al
eje del fruto. Las porciones internas y las
semillas quedan unidas a una columna central
(Ej. Cedrela lilloi, cedro tucumano).
Poricida: implica la formación de orificios
para salida de las semillas.(Ej.: Antirrhinum
majus, conejito; Papaver somniferum,
amapola)
FRUTOS SECOS INDEHISCENTES
Aquenio: fruto uniseminado, con el pericarpio
coriáceo. (ej. Ciperáceas). Cuando el pericarpio
es membranoso se lo designa utrículo.
Cariopse: el tegumento y pericarpo están
soldados entre si y a la semilla. Es el fruto
típico de las Gramíneas.
Legumbre indehiscente: a diferencia de la
forma dehiscente, permanece cerrada (ej.:
Prosopis spp., algarrobos). El maní (Arachis
hypogaea) también posee una legumbre
indehiscente, que a la madurez se entierra.
Sámara: aquenio alado (Tipuana tipu, tipa
blanca).
Esquizocarpo: fruto seco que se fracciona en
porciones monospermas (mericarpos).
Existen casos en los cuales el fruto deriva de
un ovario ínfero y reciben nombres
particulares:
Cipsela: es un aquenio procedente de un
ovario ínfero (ej. Helianthus annuus, girasol).
Cuando la cipsela va unida a un papus piloso
recibe el nombre de tricosura (ej. Taraxacum
officinale, diente de león).
Nuez: aquenio de ovario infero con pericarpio
lignificado (ej. Corylus avellana, avellano).
FRUTOS CARNOSOS
Baya: es el típico fruto carnoso, de epicarpio
membranoso, meso y endocarpo carnosos
(ej. Lycopersicon esculentum, tomate).
Hesperidio es una variante de baya con
epicarpio grlanduloso, mesocarpio corchoso y
endocarpio jugoso (ej. Citrus sinensis,
naranja).
Drupa: es otra variante de la baya,
conepicarpio membranoso, mesocarpio
carnoso, pero endocarpo leñoso (ej. Prunus
persica, durazno).
Cuando los frutos carnosos derivan de un
ovario ínfero aparechen otros tipós:
Pomo: baya derivada de un ovario ínfero
donde la mayor parte de la porción comestible
deriva del receptáculo floral (ej. Malus pumila,
manzana).
Pepónide: epicarpio lignificado soldado a la
base de los restantes ciclos florales, con meso y
endocarpo carnosos, pluriseminado (ej.
Citrullus vulgaris, sandía).
Drupa involucrada: la porción carnosa de esta
fruta está constituída por receptáculo floral,
involucro, epi y mesocarpio. El endocarpio en
cambio es leñoso (ej. Juglans regia, nogal).
FRUTOS CARNOSOS DERIVADOS DE UNA
INFLORESCENCIA
Sícono: el receptáculo es cóncavo y en su
interior se insertan los ovarios de numerosas
flores que se transforman en aquenios (ej.
Ficus carica, higo).

73 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Sorosio: el receptáculo es convexo o alargado
y sobre él se disponen bayas (ej. Morus alba,
mora; Anana comosus, ananá).
POLINIZACIÓN Y DESARROLLO DEL FRUTO
El desarrollo del fruto se inicia en
muchos casos antes de la fertilización, cuando
el polen comienza a crecer en el estigma. El
polen posee hormonas (usualmente auxinas)
que desencadenan una respuesta en los tejidos
del ovario, que empiezan a crecer y a su vez
generan sus propias hormonas, frecuentemente
etileno, que acelera la caída de los pétalos.
Muchas veces el desarrollo del fruto requiere
que se produzca la fertilización y el desarrollo
de semillas. Si las semillas no se desarrollan o
sólo unas pocas lo hacen, el fruto no crece o
queda deformado. En ciertas especies, el
crecimiento de las semillas no es necesario, y el
desarrollo del fruto se produce con
independencia de éstas. Estos frutos sin
semillas se llaman partenocárpicos (banana,
uvas de corinto, sultaninas). Los frutos
partenocárpicos suelen tener claras ventajas
comerciales.
MADURACIÓN DE LOS FRUTOS
Cuando los frutos carnosos maduran,
sufren una serie de cambios característicos,
desencadenados por otra hormona vegetal, el
gas etileno. entre los cambios causados por el
etileno están la degradación de almidón que se
transforma en azúcares, ablandamiento del
fruto causado por la degradación de las
sustancias pécticas de la laminilla media.
Se han modificado geneticamente algunas
especies cultivadas en modo tal que no
producen etileno espontaneamente. De este
modo sus frutos no maduran hasta que el
etileno es aportado artificialmente. El primer
tomate de este tipo fue liberado para el
consumo comercial en USA en 1994. Sin
embargo la calidad de estos tomates no ha sido
adecuada y no han tenido gran éxito en el
mercado.
LA DISPERSIÓN DE LOS FRUTOS
La dispersión de los frutos es
generalmente un proceso esencial para lograr
un establecimiento y propagación efectiva de
cada especie. Un factor esencial en la
dispersión es el agente dispersor. Según éste se
habla de:
anemocoria: dispersión por viento.
hidrocoria: dispersión por agua.
zoocoria: dispersión por animales.
antropocoria: dispersión por el hombre.
LA MADURACIÓN Y DISPERSIÓN DE LOS
FRUTOS Y SEMILLAS SILVESTRES EN LOS
BOSQUES DEL NOA
Las especies leñosas del NOA
presentan un comportamiento estacional, con
una mayor abundancia de fructificación durante
la estación húmeda. Pero existe una
diferenciación según el mecanismo de
dispersión. Durante la estación lluviosa
(diciembre-marzo) maduran y caen o son
consumidos preferentemente los frutos carnosos
(dispersados en especial por aves: chal-chal,
arrayan, mato). Mientras que las especies
anemócoras (tipa, jacaranda, cedro tucumano)
se dispersan durante el período seco, desde
fines de invierno a fines de primavera.
IMPORTANCIA ECONÓMICA DE LOS FRUTOS
Los frutos se encuentran entre los
productos agrícolas más importantes. Cumplen
un papel importante como fuente de
carbohidratos, aunque relativamente menor
comparado con los cereales. Botanicamente, los
cereales también son "frutos" (aquenios),
aunque las sustancias alimenticias se
encuentran principalmente en los tejidos
seminales. Aunque su valor energético por
unidad de peso en los frutos carnosos es menor
que en las semillas, su papel en la dieta es muy
importante como fuente de minerales, vitaminas
y fibra. La tabla 1 muestra una lista de los
frutos de mayor producción cultivados en el
mundo.
El fruto comercial más importante en la
provincia de Tucumán es el limón (Citrus
limonia), del que se producen más de 1.000.000
toneladas. Unos 6.500.000.000 limones cada
año.

74 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Fruta Nombre científico Familia Tipo de fruto Usos principales
Uva Vitis vinifera Vitaceae Baya Producción de vino, consumo fresco
Naranja Citrus sinensis Rutaceae Baya (hesperidio)
Producción de jugo, consumo fresco
Banana Musa spp. Musaceae Baya (clamidobaya)
Consumo fresco
Tomate Lycopersicon esculentum Solanaceae Baya Consumo fresco, enlatado, procesamiento como pasta o salsa.
Manzana Malus domestica Rosaceae Pomo Consumo fresco
Coco Cocos nucifera Palmeras Drupa Consumo fresco, deshidratado y molido
Durazno & nectarines
Prunus persica Rosaceae Drupa Consumo fresco, enlatado
Sandía Citrillus vulgaris Cucurbitaceae Pepónide Consumo fresco
Mango Mangifera indica Anacardiaceae Drupa Consumo fresco, procesamiento industrial
Pera Pyrus communis Rosaceae Pomo Consumo fresco, enlatado
Ananá Anana comosus Bromeliaceae Sorosio Consumo fresco, enlatado
Dátil Phoenix datylifera Palmeras Drupa Consumo deshidratado
Mandarina Citrus reticulata Rutaceae Baya (hesperidio)
Consumo fresco
Palta Persea americana Lauraceae Baya Consumo fresco, procesamiento como pasta
Frutilla Fragaria (varios hibridos) Rosaceae Conocarpo Consumo fresco, procesamiento como pulpa
Tabla 1. Los principales frutos comerciales consumidos por el hombre.
Figura 4.7.1. Producción de etileno. Desarrollo del fruto de frutilla desde la antesis hasta la madurez. Al
producirse la polinización se genera un pico de gas etileno que produce la caída de los pétalos. Posteriormente los
carpelos se desarrollan en aquénios, cuya producción de hormonas estimula el desarrollo del receptáculo. Cuando
el receptáculo alcanza su tamaño final se empieza a producir gas etileno que dispara una serie de transformaciones:
degradación de la clorofila, aumento de las antocianinas, reducción de los ácidos orgánicos, aumento de los
azúcares solubles, degradación de la pectina, generación de ésteres volátiles perfumados.

75 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
4.8. SEMILLA Características generales
La semilla es el órgano de las plantas con flores
que se desarrolla a partir del óvulo fecundado.
Es una etapa extremadamente importante en la
vida de un vegetal, esencial para la dispersión y
reproducción de la especie. Esencialmente una
semilla está constituida por un embrión,
sustancias de reserva, y una cubierta
protectora. La cubierta es un elemento
destinado a proteger al embrión y las reservas
de los distintos factores físicos y biológicos
hasta que se inicia la germinación. Las
sustancias de reserva permiten al embrión
desarrollarse y crecer hasta producir estructuras
fotosintéticas y alcanzar autosuficiencia. En
este sentido las semillas sufren dos tendencias
contrapuestas: por un lado mayores reservas
aseguran una mejor para la plántula cuando
inicia el desarrollo, pero por otro implicar
mayor peso y volumen que dificultan la
dispersión y aumentan la probabilidad de ser
destruidas por los diversos organismos que se
alimentan a base de semillas.
El tamaño de las semillas
Las sustancias de reservas determinan en gran
medida el tamaño y volumen alcanzado por una
semilla. Las semillas más grandes perteneces a
la familia de las palmeras que pueden alcanzar
un peso de algunos kilogramos. El ejemplo más
notable es probablemente el coco de las islas
Seycheles (Lodovicea maldivica) que puede
alcanzar 20 kg. Sin embargo la mayoría de las
semillas suelen pesar menos de un gramo. Así
por ejemplo una sola planta de amaranto
(Amaranthus sps.) puede producir unas 50.000
semillas. En el extremo inferior las semillas
pueden pesar apenas algunos microgramos,
como en el caso de las semillas de orquídeas.
Una sola cápsula de orquídea puede contener
fácilmente 1-2 millones de semillas. Existe
generalmente una relación inversa entre tamaño
y número de semillas por planta.
Importancia económica
Las semillas constituyen una fuente de energía
para numerosos organismos, principalmente
hongos, insectos y vertebrados. Entre ellos se
encuentra el hombre, que es el principal
consumidor mundial de semillas. Las semillas
de los cereales (gramíneas) y legumbres
(leguminosas) son la base de la alimentación
humana.
En la tabla 4.8.1 aparece una lista de las
principales semillas consumidas por el hombre
y de algunas de las restantes especies
alimenticias que se cultivan en escala similar a
nivel mundial.
Las partes de las semillas
1. Cubiertas seminales.
Las cubiertas seminales se originan en los
tegumentos del óvulo que sufren cambios
durante la formación del embrión y las
sustancias alimenticias. Si el óvulo tiene dos
tegumentos (primina y secundina), las capas
derivadas se denominan testa, la capa externa
usualmente más resistente, y tegmen,
comúnmente más delgada y menos resistente.
En general la dureza y resistencia de las
cubiertas están en función del medio de
dispersión de la semilla.
En muchos casos la cubierta seminal se
encuentra formando pelos, excrecencias,
ganchos, alas, etc.
En la cubierta seminal se observan
cicatrices asociadas a su origen como óvulo.
Estas cicatrices son:
La micrópila, un pequeño poro que existe ya en
el óvulo. Suele observarse como una depresión.
El hilo, que es la cicatriz dejada en la zona de
unión luego de desprenderse del funículo.
Suele ser una zona particularmente permeable
al agua.
El rafe, que es un resalto presente en el
tegumento en las semillas derivadas de óvulos
anátropos, en la zona de contacto con el
funículo.

76 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
En algunas semillas se presentan
excrecencias particulares que reciben el nombre
de arilo, ariloide y carúncula. Todas estas
estructuras suelen tener un rol muy importante
en la dispersión de las semillas, usualmente
realizada por animales.
El arilo es una excrecencia carnosa del
tegumento externo que puede presentarse
cubriendo parcial o totalmente la semilla. Según
la ubicación se puede hablar de arilo micropilar,
arilo rafeal, arilo general.
Ariloide es un término que modernamente se
está empleando para referirse a pequeñas
excrecencias del tegumento, o elaiosomas, que
son atractivos para las hormigas que intervienen
en su dispersión.
Carúncula es un tipo particular de arílo
desarrollado a partir de la zona micropilar.
Estrofiolo es otro tipo particular en el cual es
desarrollo se produce en la zona del funículo o
del rafe.
2. El embrión
En el embrión maduro se distinguen
una serie de partes:
La plúmula, que es el eje caulinar
embrionario, usualmente poco desarrollado, con
o sin pequeños primordios foliares no
expandidos. En las gramíneas la plúmula se
encuentra envuelta por una membrana que se
denomina coleoptile.
El o los cotiledones, que son las
primeras hojas, originadas en el embrión.
Suelen ser morfologicamente diferentes de los
nomófilos y adaptados a distintas funciones
dentro de la semilla: almacenamiento de
sustancias de reserva, absorción haustorial de
sustancias alimenticias almacenadas y
fotosíntesis durante las primeras etapas de la
plántula.
La radícula, que es la raíz embrional,
usualmente el primer órgano que sale de la
cubierta seminal. En el caso de las gramíneas, la
radícula se encuentra cubierta por una
membrana denominada coleorriza.
La porción de eje caulinar que se
encuentra por encima del nudo cotiledonar,
entre los cotiledones y el ápice caulinar, se
denomina epicótile. Es la porción que se elonga
cuando los cotiledones quedan dentro de las
cubiertas seminales.
La porción de eje caulinar ubicada entre
el nudo cotiledonar y el ápice radical se
denomina hipocótile. Esta porción se alarga en
las semillas que exponen sus cotiledones a la
luz.
3. Las substancias de reserva
El endosperma o albumen es el tejido
nutricio de la semilla, que se origina como
resultado de la unión de los núcleos polares del
saco embrionario con el núcleo trofogénico del
tubo polínico. Como fruto de la fusión de tres
núcleos haploides el endosperma posee un
complemento cromosómico triploide. Durante
su desarrollo el endosperma suele absorber las
sustancias presentes en el nucelo. Las semillas
que posee un endosperma desarrollado se
denominan albuminadas. Las semillas de
gramíneas son un ejemplo típico de semillas
albuminadas.
En la mayor parte de los casos aunque
el endosperma está bien desarrollado, una cierta
porción de las reservas se almacena ya en el
embrión. En otros casos el endosperma solo
tiene un desarrollo transitorio, y la mayor parte
o totalidad de las reservas alimenticias se
traslocan al embrión mismo, especialmente los
cotiledones. Se habla en esta situación de
semillas exalbuminadas. Muchas especies de
leguminosas son ejemplos característicos de
semillas exalbuminadas.
Por otro lado, existen semillas en las
cuales una buena parte de las sustancias
alimenticias se acumulan en los tejidos del
nucelo. En este caso se las denomina
perispermadas. Ejemplos típicos de semillas
perispermadas se encuentran en las
quenopodiáceas, como la quinoa (Chenopodium
quinoa).
En las gimnospermas no existe doble
fecundación como en las angiospermas. En ese
caso las sustancias nutritivas se acumulan en el
protalo haploide, y se habla de semillas
protaladas.

77 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Figura 4.8.1: Semilla de gramínea con sus
partes. Arriba: corte transversal; Abajo: vista
exterior.
Apomixis.
En la mayoría de las especies las
semillas se producen como resultado de la
fecundación del óvulo. Sin embargo en un
número bastante importante de especies se
produce la formación de semillas sin que
intervenga polen o sin que haya una unión de
gametas. Se habla en este caso de apomixis. En
la apomixis el embrión de la semilla se genera
por distintos procesos, que pueden ser:
A partir de ovocélulas no fecundadas,
pudiendo ser estas haploides o diploides. Se
habla entonces de partenogénesis.
A partir de una célula del saco embrionario
que no es la ovocélula (las antípodas o las
sinérgidas), en cuyo caso se denomina
apogamia.
A partir de células de la nucela o de los
tegumentos, que se dediferencian originando
un embrión. Se llama este tipo embrionía
adventicia. Embrionía adventicia puede dar
lugar a la formación de más de un embrión
en cada semilla, lo que se conoce como
poliembrionía adventicia. La
poliembrionía es común en los citrus (Citrus
sps.) y en el mango (Mangifera indica).
La poliembrionía puede darse también
como resultado de la fertilización de más de una
oósfera, fenómeno bastante común en las
coníferas.
Origen y formación del embrión
Las etapas tempranas de la
embriogenia (desarrollo del embrión) son muy
similares en dicotilendóneas y
monocotilendóneas. La formación del embrión
comienza con la división del zigoto fertilizado,
dentro del saco embionario del óvulo. Las
primeras divisiones son transversales con
respecto al eje longitudinal del cigoto. Con ellas
se establece una polaridad, hacia el extremo de
la chalaza se origina el embrión mismo,
mientras que hacia el extremo de la micrópila se
origina el suspensor. El suspensor es una
estructura que ancla el embrión a la micrópila.
Posteriormente se produce una
proliferación de divisiones, que dan al embrión
una forma más o menos esférica. En
organismos como Pinus (gimnosperma) o
Selanginella (pteridofita) el suspensor cumple
el rol de “empujar” el embrión en desarrollo
hacia el interior de los tejidos nutritivos
(protalo). En las plantas con flores el suspensor
tiene además un papel imprtante en la absorción
de nutrientes del endosperma y probablemente
en la síntesis de ciertas proteínas que son
utilizadas por el embrión.
Cuando recién se forma el embrión
consiste de una masa de células relativamente
indiferenciadas. Sin embargo, rápidamente se
producen cambios en la estructura interna que
resulta en el desarrollo de sistemas de tejidos.
Por divisiones periclinales se genera la
protodermis, que será la futura epidermis.
Posteriormente cambios en la vacuolización de
distintas zonas dan como resultado la
diferenciación en procambium (el origen del

78 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
sistema vascular) y el meristema basal (que
originará los restantes tejidos).
Usualmente se distinguen distintos estadíos en
el desarrollo del embrión que reciben nombres
particulares. La etapa del embrión con forma
esférica antes del desarrollo de los cotiledones
se llama etapa globular. En las dicotiledóneas
al desarrollarse los cotiledones se habla de la
etapa en forma de corazón, que en las
monocotiledóneas es de forma cilíndrica.
Luego, al alargarse el eje radical se describe
como la etapa torpedo.
En las primeras etapas del desarrollo
del embrión las divisiones celulares se producen
en toda la estructura, pero a medida que
progresa el proceso, las divisiones comienzan a
restringirse a los meristemas apicales.
Durante el desarrollo del embrión hay un
masivo pasaje de nutrientes de la planta madre a
los tejidos de la futura semilla, que determina
una acumulación de sustancias en el
perisperma, endosperma o los cotiledones.
Por último el funículo que conecta el
óvulo con los tejidos del fruto se separa, total o
parcialmente, interrumpiendo el flujo de
sustancias. El óvulo se transforma entonces en
una entidad aislada nutricionalmente, que
comienza a perder agua. Los tegumentos del
óvulo se secan y endurecen, formando una
“armadura” protectora.
Germinación y plántula
El primer signo de que la semilla está
germinando es comúnmente el alargamiento de
la radícula, que atraviesa el tegumento seminal.
El desarrollo diferencial de los distintos órganos
permite clasificar distintos tipos de
germinación. Considerando la posición y
función de los cotiledones es posible diferenciar
dos grandes tipos: 1- germinación hipógea, en
la cual el hipocótile no se alarga y los
cotiledones permanecen dentro del tegumento y
frecuentemente bajo el nivel del suelo (Vicia
faba, n.v. haba), y 2- germinación epígea, en la
cual el hipocótile se alarga, los cotiledones
emergen y frecuentemente se vuelven foliosos y
fotosintéticos (Phaseolus vulgaris, n.v. poroto).
En algunas semillas con endosperma (ej.:
Ricinus communis, n.v. tártago), mientras éste
es consumido, es elevado junto con los
cotiledones hasta que la expansión de los
mismos termina desprendiendo sus restos. En
cambio en muchas semillas endospermadas
(gramíneas como Zea mays
o liliáceas como Allium cepa, n.v. cebolla) de
las monocotiledóneas el endosperma permanece
cubierto por el tegumento, enterrado o en la
superficie del suelo, mientras que el cotiledón
va absorbiendo las sustancias nutritivas. Una
estrategia intermedia encontramos en
Peperomia (piperáceas), que mantiene un
cotiledón en contacto con el endosperma,
actuando como haustorio, mientras que el otro
es epígeo y fotosintético.
Requerimientos para la germinación
El crecimiento del embrión se frena
hasta que la semilla madura y es dispersada. El
reinicio del crecimiento del embrión o
germinación es dependiente de varios factores,
internos y externos. Entre los factores externos
o ambientales más importantes están:
disponibilidad de agua, oxigeno y temperatura.
En muchos casos también es importante la luz.
La mayor parte de las semillas maduras
están normalmente en un estado deshidratado,
con sólo 5-20% de su peso total como agua. La
germinación no es posible hasta que las
semillas absorben agua, un proceso llamado
imbibición. La capacidad de imbibición de los
coloides de la semilla es muy poderosa, a tal
punto que algunas semillas son capaces de
tomar agua directamente de la atmósfera si esta
se encuentra cercana a saturación (quinoa,
Chenopodium quinoa).
Durante las primeras etapas de la
germinación la respiración es anaeróbica, pues
las cubiertas seminales dificultan la entrada de
oxígeno. Sin embargo esto no puede continuar
indefinidamente y para avanzar en la
germinación las semillas normalmente
requieren un buen nivel de oxigeno.
Muchas especies germinan en un rango
amplio de temperaturas, sin embargo
usualmente hay un umbral por debajo del cual

79 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
no germinan, típico para cada especie: mínimo:
0-5°C; óptimo: 25-30°C; máximo: 45-48°C.
A veces, aún cuando las condiciones
externas son favorables, algunos semillas no
germinan. Se dice entonces que están
dormidas. Las causas de dormición pueden ser
muy variadas (Ej. impermeabilidad de las
cubiertas; inmadurez del embrión; sustancias
inhibidoras). La dormición es de gran
importancia para la supervivencia de muchas
especies, que pueden permanecer durante
mucho tiempo en el suelo, formando un banco
de semillas, hasta que las condiciones internas
o externas cambian, estimulando la
germinación.
Cuando una semilla no germina por
estar deshidratada, de dice que la semilla está
quiescente, y reiniciará su crecimiento cuando
se hidrate. Quiescencia es direrente de
dorminción, ya que en la segunda no se produce
la germinación aunque la condiciones sean
adecuadas.
Longevidad de las semillas
Durante las últimas etapas de desarrollo de la
semilla dentro del fruto se produce usualmente
una deshidratación marcada de los tejidos
seminales, que suele hacerse más marcada al
separarse la semilla de la planta. Es este estado,
las funciones vitales de la semilla suelen estar
reducidas a un mínimo y le permiten sobrevivir
durante períodos más o menos prolongados,
antes de iniciar la germinación. Existen datos
comprobados de semillas que han permanecido
quiescentes durante mucho tiempo, mientras
que otros reportes son dudosos (Tabla 4.8.2).
Figura 4.8.3: Germinación hipógea y epígea (tomado de Bewley & Black, 1994, modificado)

BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
82
Figura 4.8.2: Distintos tipos de semillas de dicotiledóneas con sus partes (Tomado de Dimitri y Orfila,
1987.)
Figura 4.8.4: Distintos tipos de germinación en semillas con endosperma almacenador.

83 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Nombre Vulgar Nombre Científico Familia Organo Consumido
Trigo Triticum aestivum Gramíneas semilla Maiz Zea mays Gramíneas semilla Arroz Oriza sativa Gramíneas semilla Papa Solanum tuberosum Solanáceas tubérculo Cebada Hordeum vulgare Gramíneas semilla Batata Ipomea batatas Convolvuláceas raíz tuberosa Mandioca Manihot esculenta Euforbiáceas raíz tuberosa Soja Glycine max Leguminosas semilla Vid Vitis vinifera Vitáceas fruto Avena Avena sativa Gramíneas semilla Sorgo Sorghum sudanense Gramíneas semilla
Tabla 4.8.1. Los cultivos más importantes en la alimentación humana ordenados por tonelaje producido.
Especies Localidad edad status de la
semilla
comentarios
Almacenadas en
seco
cebada Tumba de Tutankamon ca. 3350 años no viable muy carbonizada
trigo varias tumbas egipcias 3000 años o más ?
reportes de estado viable,
pero fuente de los granos y
edad nunca autenticados
trigo Tebas, Egipto 4000-5000 años no viable
algo de estructura celular
conservada, pero degradada
al rehidratar.
trigo Feyum, Egipto 6400 años no viable idem.
Canna compacta
(achera)
Ruinas de Santa Rosa de
Tastil, Argentina ca. 600 años viable
incluidas en una drupa de
nogal formado un cascabel.
Se dato la drupa con C14,
edad de las semilla
probablemente similar
Albizzia julibrissin
Colectada en China,
almacenada en el museo
británico
200 años viable en
1940
germinación iniciada
accidentalmente, plántulas
viables fueron producidas
Cassia multijuga Museo de Historia
Natural, París 158 años viable
historia desde colección hasta
siembra totalmente
documentada
Enterradas en suelo
o sumergidas en
lodo
Lupinus articus Miller Creek, Alaska >10.000 años viable
Dudoso. Edad determinada
indirectamente de datos
geológicos que podrían estar
errados
Nelumbo nucifera
(loto) Kemigawa, Japón 3000 años viable
Semillas encontradas en un
bote sumergido que se dató.
Las semillas no se dataron y
pudieron haberse depositado
más tarde.
Chenopodium album
Yacimiento
arqueológico en
Dinamarca
>1700 años viable
Estratos datados. Las semillas
no se dataron, pudieron
haberse depositado después.
Tabla 4.8.2. Un análisis de los reportes sobre longevidad de semillas en estado quiescente (tomado de Bewley &
Black, 1994).

84 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
5.1. ECORREGIONES DE
TUCUMAN Y EL NOA Alfredo Grau & Silvia Pacheco Para poder ordenar sus ideas y tratando de
entender la distribución de los seres vivos y sus
causas, los biólogos usan el concepto de
ecorregión. Una ecorregión es un territorio
geográficamente definido en el que
predominan condiciones climáticas y
geomorfológicos relativamente uniformes que
determinan la presencia de grupos de especies
vegetales característicos. Las ecorregiones son
análogas y frecuentemente idénticas al
concepto más antiguo de regiones
fitogeográficas, ya que en su origen se usaban
principalmente para los vegetales.
Es importante señalar sin embargo que esta
división en ecorregiones es solamente un
modelo que permite ordenar las ideas. Por su
parte las plantas tratan de sobrevivir en todos
los lugares a donde llegan sus semillas, muchas
veces sin respetar estos modelos teóricos e
ideales de distribución que han confeccionado
los biólogos.
En Argentina se distinguen 13 ecorregiones
terrestres: Altoandina, Puna, Monte, Chaco,
Yungas, Selva Paranaense, Espinal, Pampa,
Campos y malezales, Esteros del Iberá, Estepa
Patagónica y Bosques Patagónicos. Varias de
éstas se subdividen en subregiones. Las cinco
primeras están presentes en el Noroeste de
Argentina y cuatro en la provincia de Tucumán
(Figura 5.1.2).
Superpuesto a este sistema de ecorregiones se
encuentran los ambientes transformados por el
hombre, de los cuales los cultivos constituyen
la mayor parte. Otro factor transformador del
paisaje muy importante es la ganadería
extensiva, que con frecuencia está asociada al
uso recurrente del fuego. En Tucumán, la
provincia más densamente poblada de la
Argentina, alrededor de la mitad de la
superficie está transformada en cultivos,
campos de pastoreo, ciudades y caminos, que
en una imagen satelital puede verse como un
diseño de líneas y cuadriculados geométricos
(Figura 5.1.3).
Altoandina
Ocupa las porciones más altas de los Andes y
cordones montañosos paralelos, usualmente
por encima de 3500 m. En Tucumán está
presente en las cadenas del Aconquija, Cerro
Muñoz y las Cumbres Calchaquíes. El clima es
frío, con valores extremos que pueden ser
inferiores a -20°C, gran amplitud térmica
diaria y anual, y precipitaciones
frecuentemente en forma sólida (nieve,
granizo) que no suelen superar los 300 mm
anuales. La vegetación está dominada por
pastizales bajos particularmente del género
Festuca (Figura 5.1.4) y plantas en cojín
(Adesmia spp., Azorella compacta). En las
zonas donde se concentra agua suelen formarse
“vegas” con una cubierta densa de juncáceas,
ciperáceas y gramíneas (Figura 5.1.5).
Puna
Ocupa las altiplanicies y cuencas cerradas,
muchas de ellas conteniendo salares, en el
sector noroeste de Jujuy, Salta y Catamarca,
entre 3300 y 4500 m. No está presente en
Tucumán. Las precipitaciones oscilan entre
100 y 300 mm anuales, pero la mayor
evapotranspiración y el mayor contenido salino
de los suelos hacen que las plantas estén
sometidas a mayor estrés que la ecorregión
altoandina, resultando en el predominio de
especies diferentes. Predomina la estepa
arbustiva baja dominada por tola
(Parastrephia spp.), tolilla (Fabiana spp.) y
añagua (Adesmia spp.) (Figuras 5.1.6 y 5.1.7).
Monte

85 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Es una extensa región que en el norte de
Argentina ocupa los valles y bolsones
intermontanos con sentido predominante norte-
sur. En Tucumán ocupa el valle del río Santa
María. Las lluvias oscilan entre 80 y 300 mm
anuales, con heladas invernales y temperaturas
elevadas en verano. La vegetación dominante
es la estepa arbustiva alta con abundancia de
dominada por Zigofiláceas (Figura 5.1.8),
particularmente jarilla (Larrea divaricata y L.
cuneifolia). En algunos sectores de destacan
cardones (Trichocereus atacamensis y T.
terscheckii). En las laderas rocosas suelen
establecerse comunidades densas de
bromeliáceas (Deuterochonia y Dyckia)
(Figura 5.1.9). En los cauces de ríos
temporarios o permanentes aparecen
leguminosas arbóreas como algarrobo
(Prosopis alba y P. nigra) y arca (Acacia
visco).
Chaco Árido y Chaco Serrano
Es una vasta llanura con suave pendiente hacia
el este. Esta llanura es el resultado del relleno
sedimentaria de una gran fosa tectónica chaco-
pampeana con materiales eólicos (loess) y
materiales de origen aluvial y fluvial
provenientes de las montañas. En las montañas
bajas y zonas suavemente onduladas al este de
las serranías principales del NOA también está
presente como Chaco Serrano. La temperatura
media anual varía entre 18° y 23°. Las
precipitaciones oscilan entre 400 y 700 mm
anuales. El tipo de vegetación característico es
el bosque xerófilo (Figuras 5.1.10 y 5.1.11) de
quebracho colorado santiagueño (Schinopsis
lorentzii), quebracho serrano (S. haenkeana), y
quebracho blanco (Aspidosperma quebracho-
blanco), a los que se agregan yuchán (Ceiba
chodatii), varias cactáceas (Trichocereus
terscheckii, Stetsonia coryne, Opuntia quimilo)
y arbustos de los géneros Acacia y Capparis.
En las zonas deprimidas predominan
algarrobos (Prosopis spp.) y chañar (Geoffroea
decorticans).
En la provincia de Tucumán la cuenca de
Tapia-Trancas está ocupada en su mayor parte
por Chaco Serrano, mientras que todo el sector
Este de la provincia presentaba originalmente
Chaco Árido. Esta zona se encuentra
actualmente muy transformada en campos de
cultivo, principalmente soja, trigo, maíz, y
campos de pastoreo.
Yungas
Esta ecorregión es también conocida como
Selva tucumano-oranense o Selva tucumano-
boliviana. Es el extremo austral de una franja
de selvas y bosques de montaña que se
extienden en las laderas orientales húmedas de
los Andes desde Colombia hasta Catamarca.
La altitud oscila entre los 300 y los 3500 m de
altitud. Las precipitaciones oscilan entre 800 y
2000 milímetros anuales, con máximos a 3000
mm algunos años, que caen en un 80% en el
período diciembre-marzo. La niebla
(“precipitación horizontal”) también
contribuye aumentando significativamente la
precipitación efectiva. Suele diferenciarse en
“pisos” altitudinales:
1) Selva de Transición o Pedemontana. En
Tucumán ocupaba las llanuras en contacto con
las montañas, donde lo árboles dominantes
eran el cebil colorado (Anadenanthera
colubrina), el pacará o timbó (Enterolobium
contortisiliquum) y la tipa (Tipuana tipu).
Actualmente esta selva ha sido casi totalmente
transformada en cultivos de caña de azúcar,
limón y hortalizas. En el norte de Salta todavía
existen algunos sectores con este tipo de selva,
en este caso dominados por el palo blanco
(Calycophyllum multiflorum) y el palo amarillo
(Phyllostylon rhamnoides), que actualmente se
encuentran en proceso de transformación a
campos agrícolas.
2) Selvas de Montaña (Yungas propiamente
dichas). Ocupa las laderas de las montañas
hasta 1500-1800 m de altura. Allí se producen
las mayores precipitaciones anuales. Dominan
árboles de hoja perenne, como el laurel
(Cinnamomum porphyria) el horco molle
(Blepharocalix salicifolia). También son
importantes el cedro (Cedrela lilloi) y el nogal
criollo (Juglans australis) (Figura 5.1.12).
Bosques de Montaña, arbustales y pastizales de
neblina. Entre 1500 y hasta 3500 m aparece un
mosaico de bosques templados dominados por
aliso del cerro (Alnus acuminata) (Figura
5.1.13) o pino del cerro (Podocarpus
parlatorei), también hay bosquecitos de
queñoa (Polylepis australis), arbustos
particularmente de la familia de las compuestas
(géneros Baccharis, Eupatorium, Senecio) y
pastizales dominados por los géneros Festuca

86 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
y Stipa (Figura 5.1.14). Ciertamente los
pastizales son una formación muy distinta de
las selvas y bosques, por ello algunos autores
prefieren considerarlos una ecorregión
diferente, similar al Páramo, presente en los
Andes de Ecuador y Colombia.
El Noroeste Argentino, con su topografía muy
variada, grandes desniveles y notable
diferencia en precipitaciones, sumada a la
acción del hombre a través del ganado y el
fuego, ha dado lugar a una variedad notable
de ambientes y paisaje muy contrastantes, a
veces separados por distancias muy breves. Un
ejemplo de estos contrastes puede verse en la
figura 5.1.15. La figura 5.1.1 esquematiza la
distribución de las distintas ecorregiones en
función de la altitud (que afecta la
temperatura) y precipitaciones.
Literatura
Burkart R et al. (Editores) 1999. Eco-Regiones
de la Argentina. Administración de
Parques Nacionales. 43pp.
Brown AD et al. (Editores) 2005. La situación
ambiental argentina 2005. Fundación
Vida Silvestre Argentina. Buenos
Aires. 587 pp.
Cabrera AL 1971. Fitogeografía de la
República Argentina. Bol. Soc. Arg.
Botánica 14:1-42
Figura 5.1.1. Esquema de distribución de las ecorregiones en el NOA en función de la altitud (que
implica un gradiente amplio de temperatura), y las precipitaciones
.

87 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Figura 5.1.2. Ecorregiones y áreas transformadas en los principales cultivos en la provincia de
Tucumán.
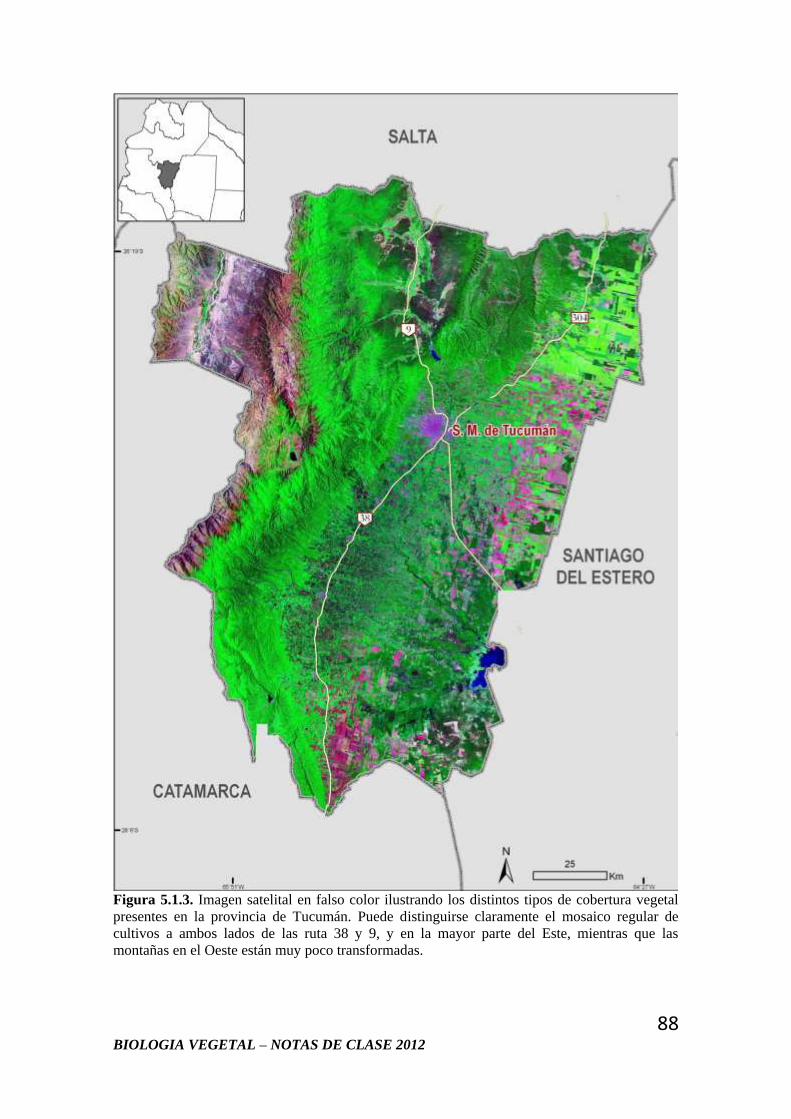
88 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Figura 5.1.3. Imagen satelital en falso color ilustrando los distintos tipos de cobertura vegetal
presentes en la provincia de Tucumán. Puede distinguirse claramente el mosaico regular de
cultivos a ambos lados de las ruta 38 y 9, y en la mayor parte del Este, mientras que las
montañas en el Oeste están muy poco transformadas.

89 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Figura 5.1.4. Ecorregión Altoandina:Pastizal de Festuca ortophylla (“iro”) formando círculos
característicos, 4400 m, cerro Muñoz, Tafí del Valle.
Figura 5.1.5. Ecorregión Altoandina:Vega en las Cumbres Calchaquíes, nacientes del arroyo
Alazán, 4400 m, Tucumán.

90 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Figura 6. Ecorregión Puneña: típica estepa arbustiva puneña dominada por Adesmia sp. y
Tetraglochyn cristatum en la zona de Tres Cruces, Jujuy, 3500m.
Figura 5.1.7. Ecorregión Puneña: Estepa arbustiva puneña dominada por Fabiana densa
(tolilla) en la zona de Yavi, Jujuy, 3700m.

91 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Figura 5.1.8. Ecorregión del Monte:Jarillal dominado por Larrea divaricata en las afueras de
Santa María, 1700 m, Catamarca.
Figura 5.1.9. Ecorregión del Monte: Cardones (Trichocereus terscheckii) y cojines de
bromeliáceas (Dyckia y Deuterochonia) en La Chilca, 1100 m, Catamarca.

92 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Figura 5.1.10. Ecorregión Chaqueña: Bosque de Schinopsis marginata con cactáceas en el
sector norte de la sierra de San Javier, 700 m, Tucumán.
Figura 5.1.11. Ecorregión Chaqueña: Bosque chaqueño con Ceiba chodatii (=Chorisia insignis,
derecha) y Cochlospermum tetraporum, río Juramento, 1000 m, Salta.

93 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Figura 5.1.12. Ecorregión Yungueña: Selva de montaña dominada por Cinnamomum porphyria
(=Phoebe porphyria), Blepharocalyx salicifolia y Cedrela lilloi, Sierra de Santa Ana, 1200 m,
Tucumán.
Figura 5.1.13. Ecorregión Yungueña:Bosques de montaña dominados por Alnus acuminata
alternados con pastizales intensamente pastoreados, El Rincón, 2400 m, Tucumán.

94 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Figura 5.1.14. Ecorregión Yungueña:Pastizales de neblina dominados por Festuca hieronymii,
Carapunco, Valle de Tafí, 2800 m, Tucumán.
Figura 5.1.15. Ecorregión Yungueña: Pastizal de neblina con cardones (Trichocereus
terscheckii, característico de ambientes chaqueños) en una ladera expuesta al NE, al lado de
selva de yungas dominada por Cedrela lilloi y Juglans australis en la ladera expuesta al SE,
cuesta de Anfama, 1500 m.

95 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
5.4. LAS PLANTAS Y EL
CAMBIO GLOBAL Las plantas y la atmósfera
Elemento o
sustancia
Proporción
(%) Comentarios
Oxigeno O2 21 oxidante, muy reactivo
O3 <0.01
Ozono, otra forma molecular del oxígeno,
filtra rayos UV nocivos en la alta atmósfera,
oxidante más fuerte que el O2 y perjudicial
en la superficie terrestre
Nitrógeno N2 78 inerte, muy poco reactivo, sólo puede ser
fijado por procariontes y descargas eléctricas
Dióxido de
carbono CO2 0.0384
Se fija por fotosíntesis en vegetales; en los
cuerpos de agua, tiene “efecto invernadero”
Agua H2O
≤1; Variable
según
temperatura
Condensado en gotitas formando nubes
refleja intensamente los rayos solares
Metano CH4 <0.001 Subproducto del metabolismo bacteriano,
muy fuerte “efecto invernadero”
Óxidos de
nitrógeno NOx <0.001
Acción tóxica sobre los vegetales, fuerte
“efecto invernadero”
Tabla 4.3.2. Composición atmosférica.
La concentración de CO2 se expresa en partes por millón (ppm), o micromoles/mol, o
en %.
Concentración de CO2: 0,0384 % = 384 µmol.mol-1
= 384 ppm
Aumento de la concentración de CO2
La concentración de CO2 atmosférico está incrementándose
Quema de combustibles fósiles
Expansión agrícola; deforestación
Aumento de la concentración de CO2 en la atmósfera, medido en el observatorio de
Mauna Loa. La fluctuación anual de invierno a verano se debe a la fijación que efectúa
la vegetación.

96 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Figura 4.3.16. La concentración de CO2 atmosférico está aumentando, desde unos 270 ppm (0,027%),
antes de la revolución industrial, hasta más de 380 (0,038%) actualmente. Este gráfico muestra los datos
obtenidos en la cumbre del volcán Mauna Loa, en la isla de Hawaii, a 4000 m de altura, que se considera
una buena referencia por no estar influida por las fuentes antrópicas de CO2.
CO2 y estomas
Respuesta de los estomas a distintos factores internos y externos
Factor Estado Respuesta
Condición hídrica Buena Apertura
Deficitaria Cierre
CO2 interno Elevado Cierre
Bajo Apertura
Ácido Absícico Elevado Cierre
Bajo Apertura
Luz Intensa Apertura
Pobre o nula Cierre

97 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
En general puede decirse que el incremento de CO2 atmosférico permite a las plantas
ahorrar agua ya que pueden absorber la misma cantidad manteniendo los estomas menos
abiertos. Aumenta la eficiencia en el uso del agua y aumenta la productividad.
Efecto morfogenético del CO2: el incremento de disminuye la densidad estomática
F. I. WOODWARD (1987) Stomatal numbers are sensitive to increases in CO2
from pre-industrial levels. Nature 327, 617 - 618
Se encontró una reducción del 40% en la densidad estomática de 8 especies de
árboles de clima templado coleccionados durante los últimos 200 años.
Esta reducción se pudo confirmar experimentalmente en algunas de las
especies cultivándolas en cámaras con una atmósfera controlada
Cambios en la vegetación terrestre
El cambio anual en superficie es pequeño, (<1%) ya que la deforestación en los trópicos
se ve compensada en buena medida por la reforestación que se produce en las zonas
templadas (y otras regiones). Sin embargo, la transformación de bosques tropicales y
subtropicales en cultivos libera gran cantidad de CO2 a la atmósfera que requiere varios
años de fijación para ser compensado.
Comparación de la deforestación en el mundo. Emisiones de carbono en Gg/año (1Gg
=1000 toneladas)
Global Tropical 970,000
Latinoamérica 460,000
Brasil Amazonas 220,000
Colombia 30,500
Argentina 20,875
México 19,066
Los frentes de deforestación en el norte de Argentina

98 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Comparación de las emisiones de CO2 de los distintos sectores productivos en la
Argentina. Nótese que la agricultura aumenta cuando se considera el CO2 equivalente
debido a la importancia del metano con gran acción invernadero.
Cambios en la cobertura boscosa de Tucumán
La cobertura boscosa de las áreas montañosas de Tucumán, donde se producen las
precipitaciones más elevadas, no ha sufrido pérdidas significativas en los últimos 35
años. Por el contrario, los bosques se han incrementado, conjuntamente con la cobertura
vegetal en general.
El gran proceso de transformación del pedemonte en áreas de cultivo, principalmente
caña y citrus, ya estaba concluido pasada la mitad del siglo XX.
En cambio la deforestación ha sido muy importante en el este de la provincia, en la zona
con bosques chaqueños secos, donde los bosques chaqueños han desaparecido. Este
proceso de deforestación continúa en las provincias de Salta y Santiago.

99 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
CO2 y efecto invernadero
Posibles líneas de acción
Reducir las emisiones de CO2
Aumentar la fijación de CO2 por parte de las plantas
¿Qué podemos hacer?
Plantar árboles
Bonos de carbono
Algunos números
16.5 toneladas CO2/persona/año (consume per capita, promedio global)
C x 3,66 = CO2
1 m3 gas = 1 kg CO2
1 l gasoil = 2,7 kg CO2

100 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
1 l nafta = 2,2 kg CO2
Lapacho (Handroanthus = Tabebuia) 30 cm Ø DAP 1380 kg CO2
12 lapachos=emisiones anuales de 1 persona
Cedro (Cedrela lilloi)60 cm Ø DAP 1570 kg CO2
Algarrobo (Prosopis spp.) 30 cm Ø DAP 1098 kg CO2
En el NOA existen grandes extensiones forestables con Prosopis. Las zonas forestables
con Cederla son menos, y se encuentran principalmente en zonas montañosas, que ya
tienen bosque en su mayor parte.
Bonos de Carbono
El mercado de compra de carbono ha crecido exponencialmente desde su creación en
1996.
Estimación del mercado mundial de carbono según el Banco Mundial
Volumen MtCO2 Millones de U$D
2005 718 10,908
2006 1,745 31,235
2007 2,983 64,035
Plantas y capa de ozono.
El ozono en las capas altas de la atmósfera tiene un efecto benéfico, filtrando los rayos
ultravioleta.
Los Clorofluorocarbonos (CFC) que se han empleado durante muchos años como gas de
refrigeración favorecen la descomposición del Ozono en la alta atmósfera, creando lo
que se conoce como el agujero de ozono.
Desde hace algunos años se han tomado medidas para limitar el uso de estos
compuestos (los CFC) y otros compuestos químicos como el bromuro de metilo
(esterilizante agrícola) y la capa de ozono está retornando lentamente a la normalidad.
El aumento de la radiación ultravioleta podría tener efectos negativos sobre los
animales.
Las plantas tienen un buen repertorio de compuestos químicos (carotenoides,
flavonoides) que filtran de manera eficiente los rayos ultravioleta, aún en los lugares
donde esta radiación es elevada.

101 BIOLOGIA VEGETAL – NOTAS DE CLASE 2012
Tendencias globales de los principales gases con efecto invernadero en los últimos años.
Efecto del ozono en la superficie terrestre
Cerca de la superficie terrestre, en la troposfera, el ozono se forma cuando la luz solar
causa reacciones químicas complejas que involucran los óxidos de nitrógeno (NOx) e
hidrocarburos volátiles, que se originan principalmente por la quema de combustibles
derivados del petróleo.
Las plantas también producen este tipo de hidrocarburos volátiles en cierta proporción.
El ozono penetra a las hojas a través de los estomas y actúa como un oxidante muy
fuerte, causando daños que pueden llegar a la necrosis foliar. Sin embargo estos
síntomas son difíciles de diferenciar de senescencia normal u otras necrosis patológicas.
El ozono tiene acción negativa en concentraciones muy bajas (ppb: partes por billón)
Las monocotiledóneas son en general más tolerantes que las dicotiledóneas a la acción
del ozono.