04 apunts genetica
TRANSCRIPT

LES LLEIS DE MENDEL Gregor Mendel (1822-1884), sacerdot austríac de l’ordre dels agustins, va establir les bases de la Genètica. Ho va fer amb l’estudi estadístic de la descendència dels cre-uaments fets amb pèsols al jardí del convent. Va estudiar caràcters senzills d’observar, després d’haver-se assegurat de treballar amb línies pures. A partir de les freqüències observades en les generacions filials va enunciar les seves lleis d’hibridació. Els seus treballs van passar desapercebuts i només amb posterioritat a la seva mort, al 1900, altres investigadors van arribar a les mateixes conclusions i la comunitat científica va poder valorar la importància real de les seves observacions. Primera llei de Mendel
Llei de la uniformitat de la 1ª generació (F1): del creuament de dues races pures per a un caràcter en surt una descendència híbrida uniforme, tant fenotípicament com genotípica. Dit d’una altra manera, del creuament d’homozigots en surt una F1 d’heterozigots.
Un homozigot per a un caràcter només pot formar un tipus de gàmeta, doncs la meiosi els forma separant dos cromosomes ho-mòlegs portadors del mateix al·lel; en l’exemple, un homozigot LL només forma gàmetes haploides L, tal com un homozigot ll només forma gàmetes haploides l. Es conclou que l’aparellament d’homozigots (línies pures) només pot donar heterozigots (hí-brids), perquè només hi ha una possible combinació de gàmetes per donar un únic tipus de zigot. La primera generació filial (F1) serà homogènia genotípicament (tots heterozigots) com fenotípi-cament (tots amb el mateix aspecte).
En el cas del color de la llavor, la F1 surt 100% groga perquè l’al·lel que determina llavor groga és dominant sobre el que determina llavor verda (L > l). En el cas que els dos al·lels presentin codominància (il·lustració de l’esquerra), els individus de la F1 són fenotípicament diferents dels dos progenitors, però iguals entre ells; per tant, també es compleix la 1ª llei de Mendel (uniformitat de la F1).
Segona llei de Mendel
Llei de la segregació dels caràcters antagònics en la 2ª generació filial: en creuar entre si els híbrids obtinguts en la 1ª generació filial, els caràcters antagònics (al·lels) se separen entre els diferents gàmetes, donant lloc a diferents fenotips en la descendència.
La meiosi permet a un heterozigot (híbrid) separar les parelles de cromosomes homòlegs per formar dos tipus de gàmetes en proporcions similars. Quan els gàmetes es combinen a l’atzar per donar la F2 aparei-xen unes proporcions genotípiques característiques: la meitat són heterozigots (com els de F1) i reaparei-xen els dos genotips homozigots (com els de P), una quarta part per a cadascun d’ells.
Les proporcions fenotíquiques co-incideixen amb les genotípiques si els al·lels són codominants. (1:2:1). Però si hi ha dominància, els heterozigots són indistingibles dels homozigots dominants; lla-vors les proporcions fenotípiques observades són 3:1.

Tercera llei de Mendel
Llei de l’herència independent dels caràcters: els diferents caràcters mendelians s’hereten indepen-dentment els uns dels altres, combinant-se a l’atzar en la descendència.
Suposem que estudiem simultàniament dos caràcters del pèsol: textura de la llavor i color de la llavor. Les característiques de la herència de tots dos caràcters s’exposen en la taula següent:
Suposem que disposem de línies pures (homozigots) pels caràcters estudiats. Triem una planta homozigota de llavor llisa i groga (LLGG) i una altra també homozigota de llavor rugosa i verda (llgg). Per tant el creuament que es vol fer amb aquesta generació paterna serà el següent: LLGG x llgg.
Òbviament cadascun dels caràcters ha de complir la 1ª i la 2ª llei de Mendel. Per tant esperem que la F1 sigui homogènia tant geno-típicament com fenotípica. Els individus de la F1 han de ser hete-rozigots per la textura de la llavor (tots Ll i de llavor llisa) i també heterozigots pel color de la llavor (tots Gg i de llavor groga). En conclusió, el 100% dels individus de la F1 seran heterozigots pels dos caràcters (LlGg).
El següent pas es creuar (o autofecundar, doncs tots són LlGg ) dos individus de la F1 per obtenir la F2. El caràcter textura de la llavor compleix la 2ª llei de Mendel, ja que es comprova que 3/4 dels pè-sols són llisos (L_) i 1/4 són rugosos (ll). També compleix la 2ª llei de Mendel el caràcter color de la llavor, doncs 3/4 dels pèsols són grocs (G_) i 1/4 són verds (gg).
El que diu la 3ª llei de Mendel és que el fet que un pèsol de la F2 tingui una determinada textura és independent del color del pèsol, doncs l’herència de tots dos caràcters és mendeliana o independent. Així, la probabilitat que un pè-sol de la F2 sigui rugós i de color groc (llG_) es pot calcu-lar com un producte de probabilitats: només cal multiplicar la probabilitat que sigui rugós (1/4) per la probabilitat que sigui groc (3/4); el resultat és 3/16.
En un dihibridisme (creuament d’heterozigots per a dos caràcters) en que tots dos gens hi hagi un al·lel dominant (L > l i G > g) les proporcions fenotípiques característiques són 9:3:3:1.
Per entendre aquesta llei cal parar atenció al funcionament de la meiosi. Dos caràcters seguiran herència mendeliana si estan en cromosomes diferents o si estan en el mateix cromosoma però prou separats (recombinació gènica).
Caràcter Al·lels Relació entre al·lels Genotips / fenotips Textura de la llavor L: llavor llisa LL i Ll: llavor llisa
l: llavor rugosa
L > l DOMINANT : recessiu ll: llavor rugosa
Color de la llavor G: llavor groga GG i Gg: llavor groga
g: llavor verda
G > g DOMINANT : recessiu gg: llavor llisa

CONCEPTES BÀSICS DE GENÈTICA Al·lel o al·lelomorf: cadascuna de les varietats existents en la població pare a un mateix gen. Codominància: situació hereditària caracteritzada pel fet que en l’heterozigot es manifesten completa-ment els dos al·lels. Fenotípicament els heterozigots (Aa) són diferents dels homozigots (AA o aa). Creuament prova: es fa entre individus homozigots recessius (aa) amb individus que presenten el fenotip dominant (A_) per determinar si aquests últims són homozigots o heterozigots estudiant la descendència. Creuament recíproc: creuament recíproc respecte a un altre és aquell en el qual els dos trets distintius que s’estudien es troben en progenitors de sexe oposat. ( ��������������������������������������������� ) Creuament retrògrad: creuament entre els mascles de la generació filial amb femelles de la generació paterna, o, també, entre les femelles de la generació filial amb els mascles de la generació paterna. Cromosoma: cadascun dels filaments, constituïts per bc-ADN, que, en nombre constant, es troben a l’interior del nucli de les cèl·lules. Els cromosomes sexuals són X i Y; la resta són autosomes. Dihibridisme: creuament en què els progenitors són heterozigots per a dos caràcters. ( ������� ���) Diploide: cèl·lula o organisme amb 2n cromosomes, és a dir, amb n parelles de cromosomes homòlegs (i per tant amb dues còpies del gen, iguals o diferents, per a un mateix caràcter). Dominància: situació hereditària caracteritzada pel fet que, en l’heterozigot, un al·lel, dit dominant, es manifesta al fenotip, alhora que emmascara o impedeix la manifestació fenotípica de l’altre, dit recessiu. Fenotip: aspecte observable (morfològic, metabòlic, etc.) que presenta un individu d’un determinat ge-notip. El terme és aplicable d’una forma global a la totalitat de l’organisme o de forma més concreta, en relació amb un caràcter puntual. El fenotip és el resultat de la interacció genotip-ambient. (����������) Gen: fragment d’ADN que actua com a unitat hereditària, determinant l’herència i expressió d’un caràcter. Generació filial: generació que resulta d’un creuament experimental. De l’encreuament dels individus de la generació paterna en resulta la 1ª generació filial o F1 ; la F1 engendra la 2ª generació filial o F2 . Genètica: branca de la biologia que estudia l’herència biològica. Genotip: constitució hereditària d’una cèl·lula o organisme ( = genoma); de forma més restringida, la constitució hereditària en un determinat locus o per a un determinat caràcter. (����) Haploide: cèl·lula o organisme amb n cromosomes, només la meitat de cromosomes presents en les cèl·lules o organismes diploides de la mateixa espècie (és portador d’un sol al·lel per a un caràcter donat). Hemizigot: cèl·lula o organisme que, tot i tenir n parelles de cromosomes, no és homozigot ni heterozigot per al caràcter estudiat, doncs el seu locus és un cromosoma sexual i només posseeix una còpia de l’al·el. ������������������������������������������������������������������������������������������������������ ����������������!���
Herència intermèdia: cas particular de la codominància, caracteritzar pel fet que l’heterozigot, a més de ser diferent dels homozigots, presenta un fenotip intermedi. �"�����������������������������������������Heterozigot: organisme portador de dos al·lels diferents del mateix gen; Mendel en deia híbrid. Per me-iosi produeix dos tipus de gàmeta. � �������#���� ��������Homozigot: organisme portador de dues còpies idèntiques del mateix gen; Mendel en deis línia pura. Per meiosi produeix un sol tipus de gàmeta. � ������#���� � Locus (pl. Loci): posició determinada que un gen ocupa en un cromosoma. Meiosi: divisió cel·lular reduccional, lligada a la reproducció sexual, en la qual, a partir d’una cèl·lula diploide (2n), es formen quatre cèl·lules filles haploides (n); bàsica per entendre les lleis de Mendel. Monohibridisme: creuament en que els progenitors són heterozigots per al caràcter estudiat. ���������� Mutació: alteració experimentada a l’atzar per l’ADN; l’individu portador de mutació és un mutant. Proporcions o freqüències fenotípiques: en un creuament, repartició quantitativa dels diferents feno-tips entre els individus de la generació filial. Proporcions o freqüències genotípiques: en un creuament, repartició quantitativa dels diferents geno-tips entre els individus de la generació filial. Recessivitat: al·lel que, en combinació amb l’al·lel dominant, no es manifesta fenotípicament.

HERÈNCIA LLIGADA AL SEXE
En l’espècie humana, la resta de mamífers i, en general, molts animals, la determinació del sexe de l’individu depèn dels anomenats cromosomes sexuals. Amb algunes excepcions notables, les femelles tenen una combinació cromosòmica XX i els mascles, XY. Ara bé, aquests cromosomes contenen gens que res tenen a veure amb el sexe però que, pel fet de tenir el seu locus en un cromosoma sexual, se-gueixen una herència diferent de la dels gens situats als autosomes o cromosomes no sexuals. Tot seguit es fan algunes precisions importants per entendre les implicacions de l’herència lligada al sexe.
• Herència lligada al sexe: la que segueixen els caràcters no sexuals però que depenen de l’expressió de gens que tenen el seu locus en cromosomes sexuals. Es parla d’herència ginàndrica si el cromosoma sexual en qüestió és l’X. Si el locus del gen està al cromosoma Y, es fa servir l’expressió herència holàndrica, si bé és molt poc freqüent, per què el cromosoma Y acostuma a ser molt petit i conté pocs gens.
• Les femelles acostumen a ser el sexe homogamètic, doncs tots els òvuls hereten un cromosoma X. En canvi els mascles són el sexe heterogamètic, perquè formen dos tipus de gàmetes, uns que re-ben el cromosoma X i altres el cromosoma Y.
• És important no oblidar que les femelles envien els seus cromosomes X tant a fills com a filles. Però els mascles fan arribar el cromosoma X només a les filles (que seran XX) i el cromosoma Y només als fills (que seran XY). Per això, quan s’estudia l’herència lligada al sexe, cal tenir molt present que un creuament pot donar resultats molt diferents dels obtinguts en el seu creuament recíproc.
• Amb alguna excepció molt puntual (gens compartits per X i Y), els mascles no poden ser ni homo ni heterozigots pels caràcters lligats al sexe, ja que només disposen d’una còpia del gen; el terme que es fa servir és hemizigosi.
• Dos casos ben coneguts d’herència lligada al sexe en l’home són el daltonisme i l’hemofília. El daltònic és la incapaç de distingir certs colors, normalment el vermell i el verd. L’hemofílic té difi-cultats més o menys severes per dur a terme la coagulació sanguínia en cas de patir qualsevol tipus d’hemorràgia. En tots dos casos es tracta d’afeccions congènites lligades al cromosoma X i causa-des per un al·lel mutant que és recessiu respecte l’al·lel normal. Aquest fet fa que la majoria d’afectats siguin homes, ja que les dones, per manifestar el problema haurien de ser homozigotes i, per tant, haver heretat la dolència tant del pare com de la mare. En canvi els homes, que són hemi-zigots, seran daltònics o hemofílics si hereten de la mare un cromosoma X amb l’al·lel defectuós.
L’exemple serveix per veure que, quan el caràc-ter estudiat està lligat al sexe, els creuament re-cíprocs poden donar resultats diferents. En el creuament de l’esquerra la femella té els ulls vermells i el mascle, blancs. En el creuament de la dreta la femella té ulls blancs i el mascle, ver-mells (creuament recíproc). Si el caràcter fos autosòmic, un creuament i el seu recíproc donarien els mateixos resultats.

DETERMINACIÓ DEL SEXE
En el curs de la història evolutiva totes les espècies han desenvolupat mecanismes que, per barreja de material ge-nètic, generen variabilitat: en els curs d’unes poques generacions apareixen un gran nombre de genomes (combi-nacions d’al·lels) que seran seleccionats en funció del seu valor biològic. Aquests mecanismes poden ser:
• Processos parasexuals: els organismes amb reproducció asexual només podrien deixar còpies idèntiques d’ells mateixos (clons), si no fos per certs mecanismes que permeten un intercanvi parcial de material ge-nètic: els plàsmids (conjugació) i els virus (transducció) són els vehicles utilitzats pels procariotes per permetre que alguns fragments d’ADN passin d’una cèl·lula a una altra.
• Processos sexuals: els organismes amb veritable reproducció sexual transmeten jocs sencers de cromoso-mes; en algun punt del seu cicle biològic tenen lloc la meiosi i la fecundació. La sexualitat està estricta-ment circumscrita al món eucariota i és una forma eficaç de generar variabilitat.
Que hi hagi reproducció sexual no implica necessàriament que hi hagi sexes. Reproducció sexual significa sim-plement que dos organismes aporten informació genètica que passarà a la descendència. Si ambdós individus for-men gàmetes equivalents, no es pot parlar de sexes diferents; tampoc si el mateix individu pot produir els dos ti-pus de gàmetes (hermafroditisme). Cal que hi hagi dos tipus de gàmetes i que un individu de l’espècie pugui pro-duir únicament un d’aquests dos tipus: el gàmeta voluminós i immòbil (òvul) és format per les femelles mentre que el gàmeta minúscul i flagel·lat (espermatozoide) és format pels mascles. Però quina és la base genètica que fa diferents als mascles de les femelles?
DETERMINACIÓ FENOTÍPICA DEL SEXE La determinació fenotípica del sexe es produeix quan el sexe de l’individu no depèn de factors genètics, sinó de factors ambientals. Ha de quedar clar, però, que l’organisme té informació genètica per a poder desenvolupar-se com a mascle o com a femella: qui decideix quin dels dos sexes potencials es desenvolupa és l’ambient.
• Els anèlids joves de l’espècie Ophryotrocha puerilis són primer del sexe masculí i es converteixen en fe-melles més endavant.
• Els cucs marins del gènere Bonellia, en la seva fase larvària, neda lliurament i, si en acabar aquesta fase no ha trobat una femella, es diposita en els fons marí i es desenvo-lupa com a femella. Ara bé, si troba una femella, s’introdueix en els conductes genitals d’aquesta i, per influència de certes hormones, es desenvolupa com a mascle, amb la única missió de produir esperma.
• Les femelles d’alguns grans rèptils (cocodrils, caimans) dipositen els ous en zones panta-noses, formant grans cúmuls. Els ous donaran lloc a individus d’un sexe o de l’altre en funció de la temperatura d’incubació. Per sobre dels 27ºC es formaran mascles i per sota, femelles.
DETERMINACIÓ GENOTÍPICA DEL SEXE Són els gens els que indueixen l’aparició de les característiques d’un sol sexe. Hi ha diferents mecanismes genè-tics per a la determinació del sexe.
• Parells de cromosomes sexuals: una gran majoria d’animals vertebrats i invertebrats són diploides i tenen un parell de cromosomes sexuals, normalment iguals en les femelles (XX) i diferents en els mascles (XY). Les femelles produeixen, per meiosi, gàmetes porta-dors d’un cromosoma X (sexe homogamètic), però els mascles produeixen dos tipus d’espermatozoides (sexe heterogamètic) portadors, els uns d’un cromosoma X, que en fe-cundar l’òvul originaran femelles, i els altres, d’un cromosoma Y, que en fecundar l’òvul, donaran mascles.
* En aus i papallones el sexe homogamètic és el masculí i l’heterogamètic, el femení. * En els ortòpters (saltamartins) les femelles són XX i els mascles XO (manca el cromosoma Y).
• Haplodiploidia: són exemples coneguts formigues i abelles. El sexe depèn de la dotació cromosòmica: les femelles són diploides (fusió d’un òvul i un espermatozoide) i els mas-cles, haploides (partenogènesi:desenvolupament d’un òvul no fecundat).
• Determinació gràcies a un sol gen: en alguns himenòpters i dípters no hi ha cromosomes sexuals. L’herència del sexe depèn d’un gen amb dos al·lels; les femelles són homozigotes i els mascles, heterozigots.

HERÈNCIA INFLUENCIADA PEL SEXE
Existeixen caràcters, com ara la calvície en l’espècie humana i la presència o absència de banyes en algunes races ovines, que venen determinats per gens situats en regions homòlogues dels cromosomes sexuals o bé per gens au-tosòmics, però amb una manifestació fenotípica que està condicionada pel sexe de l’inidividu. Així la calvície he-reditària en l’espècie humana depèn d’un gen autosòmic i és causada per un al·lel que és dominant en l’home però recessiu en la dona (el mateix genotip dóna fenotips diferents en funció del sexe de l’individu).
AL·LELOMORFISME MÚLTIPLE Sovint els problemes de genètica plantegen situacions excessivament simplistes, amb només dos al·lels diferents per a cada gen. Però el pas del temps propicia que qualsevol dels al·lels existents experimenti més mutacions puntuals que comporten l’aparició de nous al·lels. Per això és habitual que en una població d’individus existeixin diversos al·lels per a un mateix gen. El fenomen rep el nom d’al·lelomorfisme múltiple i el conjunt d’al·lels que ocupen el mateix locus constitueix una sèrie al·lèlica.
Un cas d’al·lelomorfisme múltiple ben conegut és el del gen que determina els grups sangui-nis en l’espècie humana (sistema ABO). Tres al·lels diferents (IA, IB , i ) es combinen en ho-mozigosi o en heterozigosi per a determinar un grup sanguini (A, B AB o O). Els al·lels IA i IB són codominants entre ells i tots dos són dominants respecte l’al·lel i , que és recessiu.
AL·LELS LETALS En certs casos les mutacions que es produeixen donen lloc a al·lels que, per la raó que sigui, fan que l’individu portador no sigui viable, que vol dir que provoquen la mort en el període prenatal o també en el postnatal però abans de que l’individu ar-ribi a la maduresa i es pugui reproduir. Hom parla d’al·lels letals. Un al·lel letal dominant mai pot ser heretat, doncs l’individu portador no arribarà a la maduresa i no podrà deixar descendència. Els al·lels letals dominants s’originen per muta-ció de l’al·lel normal i són eliminats en la mateixa generació en la que apareixen. Pel contrari, els al·lels letals recessius que-den emmascarats en combinació heterozigòtica amb l’al·lel normal. Només es manifesten en un creuament entre heterozigots portadors; llavors, la quarta part de la descendència serà homozigòtica recessiva i morirà.
INTERACCIÓ GÈNICA És un error habitual creure que sempre un sol gen controla o determina un sol caràcter. En molts casos no és així. De vegades l’expressió d’un gen (a través de la transcripció i de la traducció) té més d’un efecte sobre el fenotip; en conseqüència un gen pot controlar més d’un caràcter. També es pot donar el cas que una característica vingui determinada per l’acció conjunta de dos o més gens, fet que es coneix amb el nom d’interacció gènica.
Un model teòric pot servir per entendre el concepte d’interacció gènica. En general, qualsevol procés metabòlic es duu a terme a través d’una ca-dena de reaccions, cadascuna d’elles catalitzada per un enzim diferent. Suposem que una substància precursora P pot ser transformada successi-vament en A, B i finalment C gràcies a la catàlisi dels enzims e1, e2 i e3, tal com indica l’esquema. Però cadascun d’aquests enzims és el resul-tat de l’expressió d’uns gens, representats per g1, g2 i g3.
Per tal que el procés metabòlic aconsegueixi l’objectiu de transformar el precursor P en el producte final C són necessaris els tres enzims funcio-nals. Suposem que un organisme és portador de còpies defectuoses del gen g3. No podrà sintetitzar l’enzim e3 i la via meta-bòlica querarà interrompuda en l’últim pas: no s’obté el producte final C. (en tot cas s’acumula l’intermediari B).
Però que passarà si l’organisme porta còpies defectuoses del gen g2 ? Que no disposa d’una versió funcional de l’enzim e2 , sent incapaç de transformar l’intermediari A en B . Però les repercussions van més enllà: la manca de B deixa a l’enzim e3 sense el seu substrat, sent impossible la producció de C. Podem deduir conseqüències similars per a la presència de còpies de-fectuoses del gen g1. La absència de qualsevol dels tres enzims fa impossible la síntesi de C.
Podem concloure que la capacitat per sintetitzar la substància C depèn de tres gens (interacció gènica) i que la informació defectuosa del gens g1 i g2 anul·la o deixa sense efecte al gen g3. El gen que pot anul·lar l’efecte d’un altre gen s’anomena gen epistàtic ( ≠ dominant); el gen que pot ser anul·lat s’anomena gen hipostàtic (≠ recessiu).
LA HERÈNCIA NO NUCLEAR No hem d’oblidar que els plastos i els mitocondris són orgànuls semiautònoms que posseeixen material genètic propi. Aquest ADN no nuclear conté informació que també serà transmesa a la descendència. Ara bé, tant en animals com en vegetals, els mitocondris i els plastos són transmesos únicament pel gàmeta femení (de l’espermatozoide, només el nucli haploide s’incorpora al futur zigot).
Genotip Fenotip (grup sanguini)
IA IA A
IA i A
IB IB B
IB i B
IA IB AB
i i O
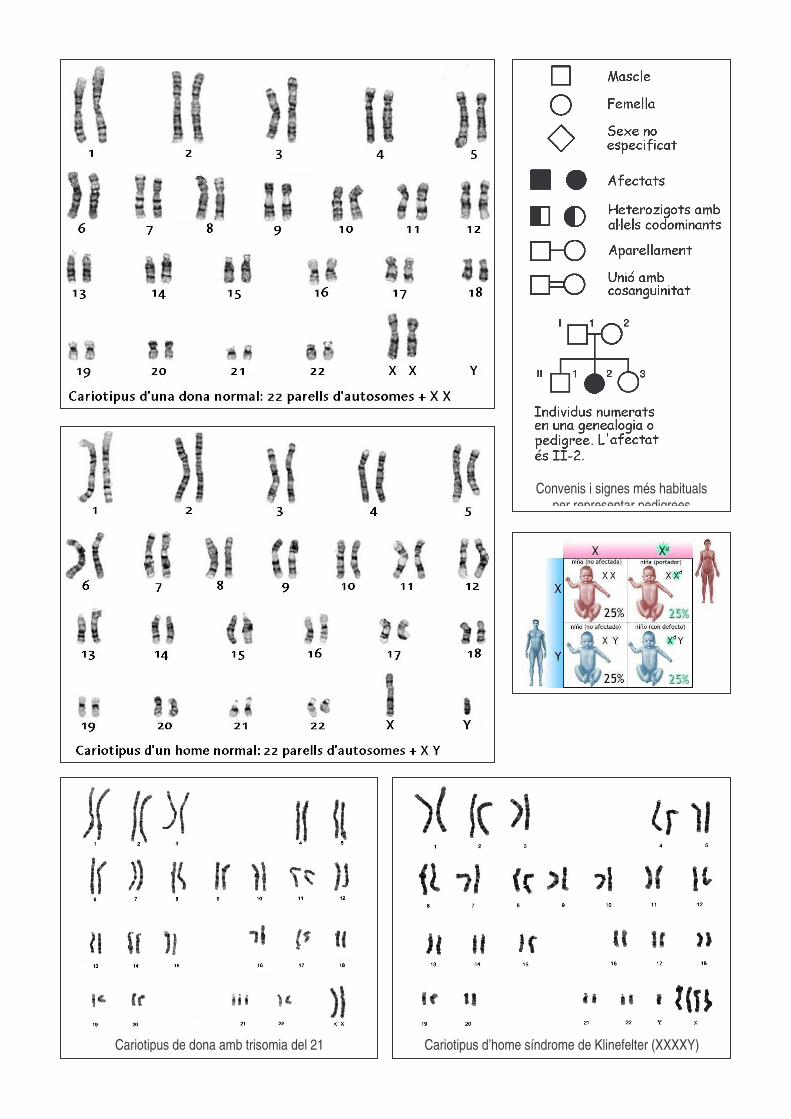
Cariotipus de dona amb trisomia del 21
Cariotipus d’home síndrome de Klinefelter (XXXXY)
Convenis i signes més habituals per representar pedigrees
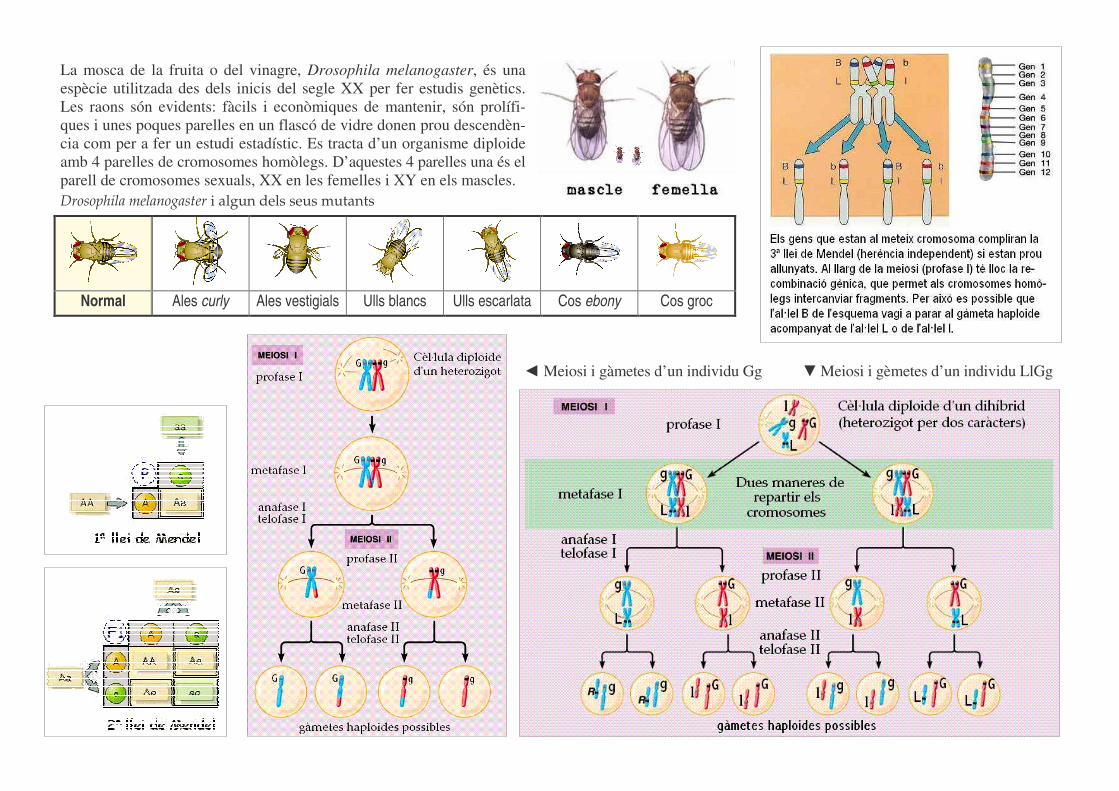
La mosca de la fruita o del vinagre, Drosophila melanogaster, és una espècie utilitzada des dels inicis del segle XX per fer estudis genètics. Les raons són evidents: fàcils i econòmiques de mantenir, són prolífi-ques i unes poques parelles en un flascó de vidre donen prou descendèn-cia com per a fer un estudi estadístic. Es tracta d’un organisme diploide amb 4 parelles de cromosomes homòlegs. D’aquestes 4 parelles una és el parell de cromosomes sexuals, XX en les femelles i XY en els mascles. Drosophila melanogaster i algun dels seus mutants
Normal Ales curly Ales vestigials Ulls blancs Ulls escarlata Cos ebony Cos groc
� Meiosi i gàmetes d’un individu Gg � Meiosi i gèmetes d’un individu LlGg