- 39 - capitulo cuatro: familia … · el género meloldogyne se conoce comúnmente bajo el nombre...
TRANSCRIPT

- 39 -
CAPITULO CUATRO: FAMILIA NETERCnJERIDAE - teNERO MELOIDOGYNE
1. GENERALIDADES
La familia Heteroderidae quizá es una de las más Importantes de la superfami
lla Tylencholdea, pues a ella pertenecen lofl géneros Meloi4ogyne y fteterodera.i
amplia distribución en muchas zonas agrícolas.
Hasta 1967, se consideraba solamente una suiífamilla - Heteroderinae - dentro
de la familia Heteroderidae. La única modificación sugerida en ese entonces
fué la adición de Hypsoperine y Meloidoderita descritos por ves primera en
1964 y 1966, respectivamente (1). Posteriormente, se hizo necesaria una revi
sión de esta familia con la cual quedaron distribuidos siete géneros en dos
subfamilias, Heteroderinae y Meloidogynlnae. Los parámetros que se tuvieron
en cuenta fueron^a_go8lclón^lel^^oro excretor, las estriaclones del cuerpo y
la posición de la^vulva. (Cuadro No. 7).
La última revisión de la familia Heteroderidae data del aflo de 1972 en la cual
Wouts (51) justificó la elevación de la subfamilia Meloidogynlnae a familia Me
loidogynldae, la que estaría constituida por Meloldogyne. Hypsoperlnae y Meloi
doderita. siendo este último género trasladado de la subfamilia Heteroderinae.
Igualmente, propuso una nueva subfamilia - Ataloderinae-en la familia Heterode
rinae.
Las características más sobresalientes de la familia Heteroderidae son las si
guientes: Pronunciado dimorfismo sexual, hembras endo o semiendoparásitas, sa
ccloformes o alimonadas y didélficas; machos mono o diórquicos, con espícula y
gubemáculo pero sin bursa.

40 -
2. GENERO MELOIDOGYNE
El género Meloldogyne se conoce comúnmente bajo el nombre del nemátodo del nu
do radical debido a que Induce en sus hospederos a la formación de abultamlen-
tos radiculares. Comu fué el primero en describirlo en 1879 y lo denominó An-
guillula gK)rionl (42). • Sin embargo,' se emplearon muchos sinónimos hasta 1932,
aflo después del cual se modificó el nonibre a Heterodera mar ion 1 (Cornu 1879)
Goodey, 1932. Los sinónimos más usados fueron:
Heterodera marloni (Comu 1879) Goodey 1932.
Sin. Anguillula marloni Comu 1879.
Heterodera radlclcola (Greef 1872) Muller 1884.
Anguillula arenarla Neal 1889.
Ansjulllula vlalae Lavergne 1901-
Heterodera javanica Treub 1885.
Tylenchus arenarlus Cobb 1890.
Meloldogyne exigua Goldl 1887.
Oxyurlus incógnita Kofoid y Whlte 1919.
Caconema radlclcola (Greef 1892) Cobb 1924.
Hubo de esperarse hasta el año de 1949 cuando Chitwood, al revlaar la clasifi
cación de los nemátodos del nudo radical, vio la necesidad de establecer un nue
vo género que estuviera representado por aquellos nemátodos, pues en su opinión,
tenían características que les distinguían de las formas de Heterodera. Con ba
se en esa consideración, modificó el nonibre genérico a Meloldosyne (18, 42).
3. BIOLOGÍA
Los huevos puestos por las heníbras de Meloldogyne son ovalados y algo alargados*

41
s a 1 o
to rt
1
a 1
a %
o
s I *
8 o
a n
o « OQ
g H o
I
Ct » 1
&
I y»
A
IB O O H
(a 'P H o n 3 9 I» <D C <• (D r» n a n rt )-• © H- « H» M o >i O a n H- H* H* (D o e o a O O 1 ^ o o rt » B» H - W
a o «< o> rfi (D n 0 a C4 I «> (D
2 •*
B» S •- o » »T3 f t •
O . O l i «o
•o rt os O í »
O O »1
t « 1
a 9 (O a n a o A o I -• I o
o tt a a • »-»|rt
o- < n c ro • -
S •o » o r t r r »
&3 o H*
Sg 9 t -O • H» (9 CD
O 1
ca < O C cr t -1 < A »
•o r t 1 O
S3 c e-n S 1 •-*
If/
s
8
g
a Ql
^ \ . T N ^ •fl o O
C
tO
c
< .c
g-o
3
o tt O
fo m n a
i
á n o < o o c
• - O" 9 (-« : 3 ° S • p» roma* to t-* 3 c » a 9 n f h* a I* e» a H-tt o* » a o H> >-• o> a o • » A g < > 9 » c 9
•-* O © • I r -
^§2. a* »-» »-• < 9 n < 0» o o S >- 9
tt o 9 . o o • O" r t • M A a*
A rt c
ida e
nal. 1
lab tero
d
© »*|?> 9
3°
rt © tt
1> s 1.*
1 rt s
ft
o H-<B >.* . . . .
o a a ©
3 1
í M
•n
«rt a*
1 rt »

- 42 -
Se encuentran envueltos por una membrana - la matriz, saco de huevo o masa ge
latinosa - en la parte extema y posterior del animal. La fecundidad de una
hembra depende de muchos factores pero sobretodo de la planta (4), ya que en
una planta no hospedera puede poner alrededor de 10 huevos. Sin embargo, se
cree que una hembra pone en promedio de 200-500 huevos (42).
En un principio, se creyó que la masa gelatinosa era una secreción del útero
pero Maggentl y Alien (25) demostraron que más bien era utia secreción de 6
glándulas rectales, la cual se efectúa o se inicia por la apertura anal, para
después rodear la vulva. Esas glándulas se hacen visibles a partir del segun
do estado larvario.
Varios factores pueden estar relacionados con la expulsión de los huevos. Mag
genti y Alien (25) sugieren que un músculo ano-dorsal controla este proceso
mientras que Blrd y Rogers (9), a la luz de estudios electrónicos, lo relacio
nan con las estructuras multivesiculares presentes en el citoplasma de las
glándulas, por desembocar al exterior por medio de canales. Las estructuras
citoplásmlcas son en gran parte a base de proteínas y ácido mucopolisacárldo.
La embriología de MeloídoRyne es similar a la de otros nemátodos. En M. Java-
nica, la escisión es casi igual y asincrónica. Los cuatro primeros blastóme-
ros son distribuidos a lo largo del eje más largo del huevo, lo Cual parece ser
una característica de los Tylenchlda. El endodermo y mesodermo se originan del
rudimento común - el blastómeto Em - en el estado de cuatro blastómeros y son
representados por blastómetos independientes É y M en el estado de 7 blastóme
ros (3).
La larva de Meloldogyne en primer estado es delgado, se encuentra arrollada den

43 -
-'7>>.(.V^"! k>-••:;><• í t - y <
\^
Fig, 15. Meloidosyne areaarí.i t ha me a i , Iniciación de la deposición de la matri?: a través de la apertura anal, B. La matriz de.spués de dos horas, C. La matriz después de tres horas. ü. La matriz después de cuatro horas., (25),
Fig. 15. A. ilembra joven de Meloidos;yne inqóguita acrita mostrando la posición de las glándulas secretoras de la matriz. B. Hembra madura del mismo neraáto <\.) ilustrando cuatro de las se ij glándiilas, (25)
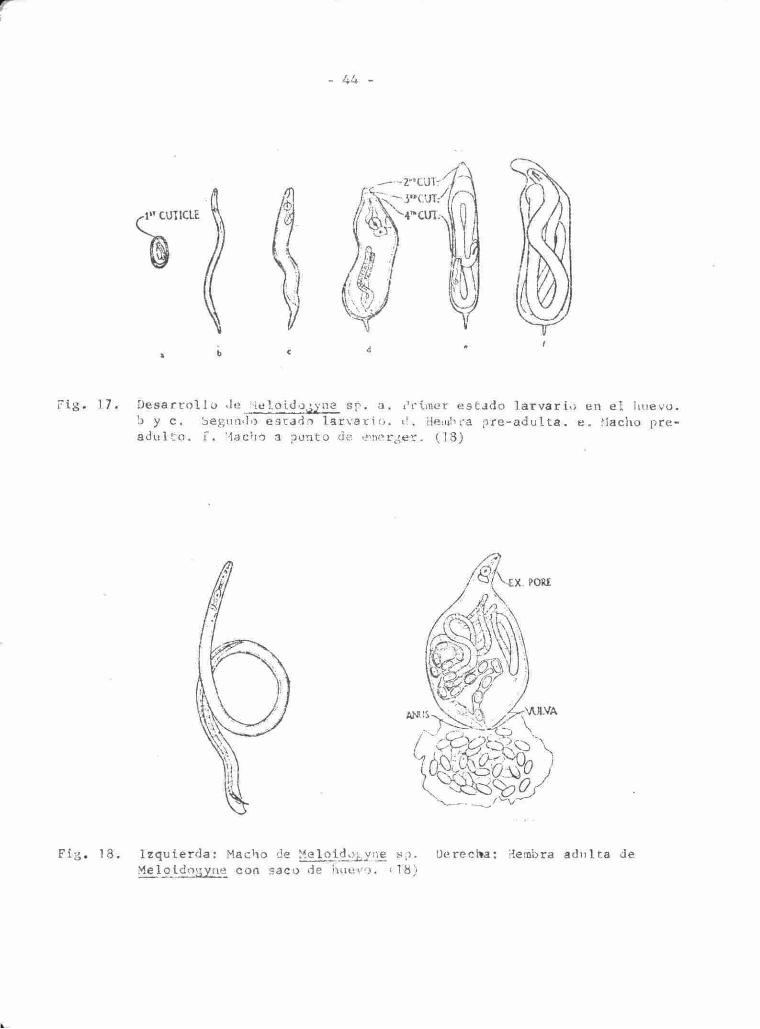
44 -
1" CUTICLE
Fig . 17. D e s a r r o l l o de Meloido,¿¿^ne sp . a. b y c . Segundo es tado l a r v a r i o a d u l t o , f, !'lacho a punto de e^nerger. (18)
i^rlmer e s t ado l a r v a r i o en el huevo, í. Hembra p r e - a d u l t a . e . Hacho pre-
y=rj-
Fig. 18. Izquierda: Macho de Maloido^yne sp. Uerecl»a: Hembra adulta de Melotdo;;yne con saco de huevo. il8)

- 45
tro del huevo y muda sólo una vez dentro de él. Posee bastante bien dlferen'?
dables el estilete, el metacorpus, el poro excretor y.el Intestino.
Según Blrd y Rogers (8), la cutícula de las larvas de M. javanlca posee tres
capas que no están separadas por membranas: una capa cortical extema osmofí-
lica, una cortical interna y otra fibrosa. La única capa que se muda es la
cortical extema y las demás, a pesar de ser secretadas por la hipodermis, se
rían reabsorbidas.
Después de la eclosión, la larva en segundo estado o larva preparáslta (L2) se
orienta hacia las raíces en busca de alimentos, lo cual se manifiesta por la
formación de las células gigantes. A pesar de que existen muchas controversias
acerca de la formación de las células gigantes, se suela explicar la mediante
dos hipótesis: 1 - Turgencia y digestión en la pared celular por medio de en
zimas hldrolíticas y pectínicas. En este caso, solamente habría fusión de las
células adyacentes al sitio de alimentación. 2 - Mitosis sin cltocineaiá;».
(49).
En los estudios de Balasubramanian y San^asvami (5), las células gigantes esta
ban localizadas en la corteza y a veces en el perlciclo. En cafla de azúcar,
estaban siempre conectadas con la endodermls.
El número de células que compone una célula gigante varía de planta a planta.
Podría ser más de 7 en cafla o 4 ^a tabaco y menos de 4 en Abelmoschus esculen-
tus. Así mismo, varía el número de células gigantes por planta::en tomate
es de 20, en tabaco 14, jsn cafla 11 y en Solanum melongena, 13. (5).
La etapa posterior es la formación de nudos o agallas, los cuales serían un úl-

- 46 -
tima instancia una reacción de laá plantas a los reguladores de crecimiento
tales como auxinas, ácido Indolacétlco, indol bbutírico y trlptófano. Hay é v i
denclas bastante sólidas de su participación en este proceso, pues hasti el m£
sentó, siempre se ha detectado mayor contenido de aminoácidos, aminas y auxi
nas en las raíces con agallas (23, 31, 36, 49).
La discrepancia entre los nemátólogos eh la procedencia de les reguladores'.
Algunos investigadores opinan que el trlptófano puede ser un producto de des
composición de las proteínas de las plantas o del complejo proteína-ácido indo
lacétlco. Sería probable también que los nemátodos lo Inyectasen a las raíces;
pero Setty y Wheeler (36) lo ven poco factible por la baja cantidad de auxinas • ' ^ . . • • • . . . . ' . ' • ' • ' ' '
que contienen los nemátodos. Las hipótesis más sólidas consideran la proce
dencia de los reguladores de crecimiento, principalmente la del ácido Indol
acético, mediante dos planteamientos (36):
1. El rompimiento del complejo lAA - proteínas mediante las enzimas proteo-
lítlcas del nemátodo y
2. Conversión del trlptófano a ácido indolacétlco mediante enzimas específj.
cas., • -•. . • " . . • ' •
Posterior a ello, la L2 empieza a hincharse y a volverse sedentaria, lo cual
se culmina al alcanzar el nemátodo el estado adulto. Trudglll (47) cree que
la durapión de la alimentación influye sobre la muda de L2, pues para ello fué
necesaria una alimentación durante 9 días a 22''C.
En muchas plantas hospederas de Meloldogyne. la corteza, la epidermis y el per¿
ciclo se ven muy afectados, formándose grandes cavidades y no así los haces
vasculares que presentan un daflo mucha más leve (5). Siddiqui y Taylor (39)

- 47 -
Cuadro No. 8 , Producción de auxina por los patógenos que inducen a desordenes
del crecimiento, A. Patógenos producen auxinas a partir de aminoácido. B.
Esquema para la producción de lAA por los patógenos: línea sólida; procesos
confirmados en experimentos con Taphrina; líneas en puntillas; procesos presu
midos con Taphrina. Estos ciclos pueden ocurrir también eh los nemátodos. (49)
A - Aminoácidos .
B - Acido antranílico
* ? índole
Auxinas
Trlptófano
1 Nemátodos
I
Insectos Femllanalina Trlptamlna Acido Índole pirúvlco Indoleacetonltrllo
\
\ Histidina
H-4IAE
I—fAN
lAA

- 48 - '
-Tabla NO. 1 . Nanomoles de aaxinád-'eh és t tac tos de pesos secos equivalentes de agallas
de plantas infestadas por Meloldogyne-spp." (49)
C O L E S R E k O L A t í H A T O M A T E
Auxina M. h. M. J[. M. i M. h. M.^:. M.i. M. h. M. ±. M.i.
lAA .
lAN
lA-E
IBA
TOTAL
34
255
39
0
328
76
238
0
0
314
6
73
8
350
437
5
50
5
0
60
27
4
0
0
31
0
0
0
Í65
165
35
1600
80
0
1715
150
30
0
0
180
0
0
0
2450
2450
lAA = Acido indole-3-acético; lAN = Indole-3-acetonitrilo; lAE = Indole-3-ácido aciéti-
co etil ester; IBA = Acido indole-3-butírico.
M, h = M. hapla; M, J, - M. javanlca; M. 1̂ . = M. incógnita.

49
CAMBIOS EN EL PARÁSITO ESTIMULADOS POR EL HOSPEDERO
Eclosión. Movimiento en el suelo
(atracción). Activación de las
glándulas ventrales. Penetración
a la corteza.
Activación de la glándula dorsal
Establecimiento en el sitio de a-
limentaclón. La L2 empieza a hln
charse y a volverse sedentaria.
M,
M3L.
M4.
L3
Adultos
Machos abandonan la raíz.
Secreción de la masa de huevo por
las glándulas rectales.
Ovoposición
DIAS
-5
O
10
15
20
25
30
CAMBIOS EN EL HOSPEDERO ESTIMULADOS POR EL PARÁSITO
Inicio de la hipertrofia corti
cal.
Hinchazón de les células vasou
lares cerca a la cabeza de la
larva.
Mitosis simultánea de los nú
cleos de las células gigantes.
Aumento de tamaño de las célu
las gigantes.
Células gigantes alcanzan su ta
maflo máximo.
Aumento en densidad del citopla¿
ma de las células gigantes. Cé
lulas gigantes del macho se de
sintegran.
Células gigantes muy bien desa
rrolladas.
Células gigantes empiezan a de
sintegrarse.
Cuadro No. 9, Interacciones en el ciclo de vida de Meloido^yne spp. Tomado de Rohde (34).

- 50 -
observaron que las L2 de M. naasl entran en las raíces de avena-a las 24 horas
de la inoculación, coincidiendo así con la hinchazón de la punta de la raíz.
A los 4-6 días, las agallas están bien desarrolladas y su aumento en tamaflo
cesa a los 12-15 días de la inoculación. Describieron varios tipos de agallas:
Agallas terminales circulares o en forma de gancho, agallas periformes sobre
las raíces laterales y agallas Intercaladas en forma de huso.
De acuerdo con Stoyanov (40), no siempre es posible encontrar especímenes de
Meloldogyne en raíces de plátano, debido a que no se producen agallas pronun
ciadas. Estas se forman principo.lmenta en las raicillas delgadas y en los ex
tremos de las zonas de crecimiento de las raíces.
Aunque Rohde (34) considera que el ciclo de vida de Meloldogyne puede durar
35 días, Grlffin y Jorgenson (22) encontraron que el de M. hapla a 25''C dura
7 semanas en papa.
Los machos de Meloldogyne no son abundantes y su presencia está relacionada
con varios factores, tales como el estado nutricional de la planta, la calidad
de los alimentos y la densidad de población del nemátodo. De A. Santos (13)
opina que a mayor número de larvas penetradas en la raíz, mayor era la rela
ción macho: hembra de M. arenaria.
Davlde y Trlantaphyllou (12) estudiaron el efecto de las aplicaciones follares
de- hidrazida, maléica sobre la diferenciación sexual de M. javanlca y Mv incóg
nita en tomate y tabaco. Encontraron una relación positiva entre este ccmpue^
to y la presencia de machos, si bien hubo diferencias entre dosis de una parte
y tiempo de aplicación por otra. El mayor porcentaje de machos se' observó en
tabaco y tomate asperjados a los 6 días de la inoculación con 6 y 12-25 mg. de

- 51 -
hidrazida malélca por planta respectivamente. Mas tarde, los mismos investi
gadores (11) corroboraron parte de esos resultados agregando que las altas dó
sis de hidrazida malélca inducen la formación de hembras Intersexuadas de M.
javanlca. En síntesis, parece que este producto Inhibe el desarrollo de las
larvas y favorece el reverso sexual<
Estos dos factores que podríamos llamar ambientales, junto con la información
genética determinan el modo de reproducción de Meloidoijyne. La fertilización
cruzada - exclusiva al menos en M. carollnensls - puede ser considerada como
la condición primitiva de reproducción de la cual se derivaría la partenogé-
nesis. Dentro de este grupo, las especies partenogenéticas meióticas son con
slderadas toas primitivas que las partenogenéticas mitótlcas (45).
A la luz de esos planteamientos y tan^lén debido a su estrecho rango de hosi>e
deros, se considera M. carollnensls como la forma ancestral de todas las espe
cies de Meloldogyne. Pasando por la raza A de M. hapla que se reproduce tan
to por fertilización cruzada como por fiartenogénesis melótlca, se llegaría a
las formas partenogenétlcas mitótlcas tales como M. aaasi y M. ̂ raminlcola.
M. arenarla. M. incógnita y M. javanlca que serían las especies más evolucio
nadas de este género (45). La partenogénesis parece predominar en M. hapla y
M. thamesi (13).
Se considera que el número básico de cromosomas es de n > 18 como en M. naasl
y M. gramlntcola. Las demás, por ser formas derivadas, tendrían n > 17, 16 y
15 (M. hapla). 54 cromosomas (M. arenarla). 40-44 (M. incógnita). 43-48 (M. ja
vanlca) .

- 52 -
4.. MOREOLOGL^ ,
Hembra: Cuerpo blanco, esférico, periforme. Cutícula con finas estrías trans
versales. Vulva y ano terminales, rodeados por estrías circulares que consti
tuyen los patrones perlneales para el diagnóstico. Reglón del labio con 6 la
bios pequeños. Estilete más delgado que en el macho o en las larvas. Metacor
pus muy bien desarrollado, seguido por un istmo corto. Poro excretor anterior
al met-acorpuo, 12-27 estrías detrás de la cabeza. Didélficas, ovarios enrosca
dos. El sistema reproductivo abarca ua gran porcentaje de la longitud del cuer
po. (2, 16, 18, 42).
Macho: Cilindrico. Terminal redoivieado, sin bursa. Estilete bien desarrollado.
Mono o diórquico. Espícula casi terminal. Poro excretor cerca del anillo ner
vioso. Labios laterales prominentes. (Figura 19).
5. IDENTIFICACIÓN DE ESPECIES
Se conocen hoy en día unas 32 especies de Meloido.'̂ yce de las cuales al menos 8
son muy importantes a causa de su distribución y gama de hospederos. Existen
varios métodos para identificar las espeeles de Meloidogyne. siendo el más usa
do el' patrón cuticular de la región que rodea la vulva ,y el ano llamado comqn-
mente patrón perlneal.
Sus estructuras más Importantes están señaladas en la figura 20. El patrón pe-
rineal;contiene dos sectores: el dorsal y el ventral, los cuales están bien djL
ferenciados i9edlante urnas líneas llamadas líneas laterales. Estas se unen en
el terminal de la cola. Cada sector se compone de estrías y el conjunto de las
estrías dorsales se llama arco. (41).

<TT^
»,<:
«¡'i;
r̂
v - - I
o ' 25y
;; r:-
' '^2^'y
' O ' !
- 53
V . 1
CJ
> o
o
F t:
•10
• o
V^. A r r i l i a : wL'üiiios .le >:i\ -, l i - ^l"^ 1^ t i c o l a . A, :; v po l a t e r a l y j - c i r t e nob t ll. P a r t e a n t e r i o r y ¡.>.is A ' ' a ' ; > : D Í O M Í O de l a p a i
hr.a de '•'.••I J i d o ^ y a e ¿'¿am
r..s
' ' ' ' ^ • : - ' ' y -̂ [•
>;u:'io y da ' lan l a i v a '.ic c:. ..'^,rt;e a n t e r i o r , c-iri e r i o r de MO r t acno . i; y f e r i . o r de a n a l a r v a , í e a o t p . r i o r de r.:,a ]<•••> i a l e ó l a • !.'?'])
¡ / • • • • • • • y
:# ' •>^'-p¿.'
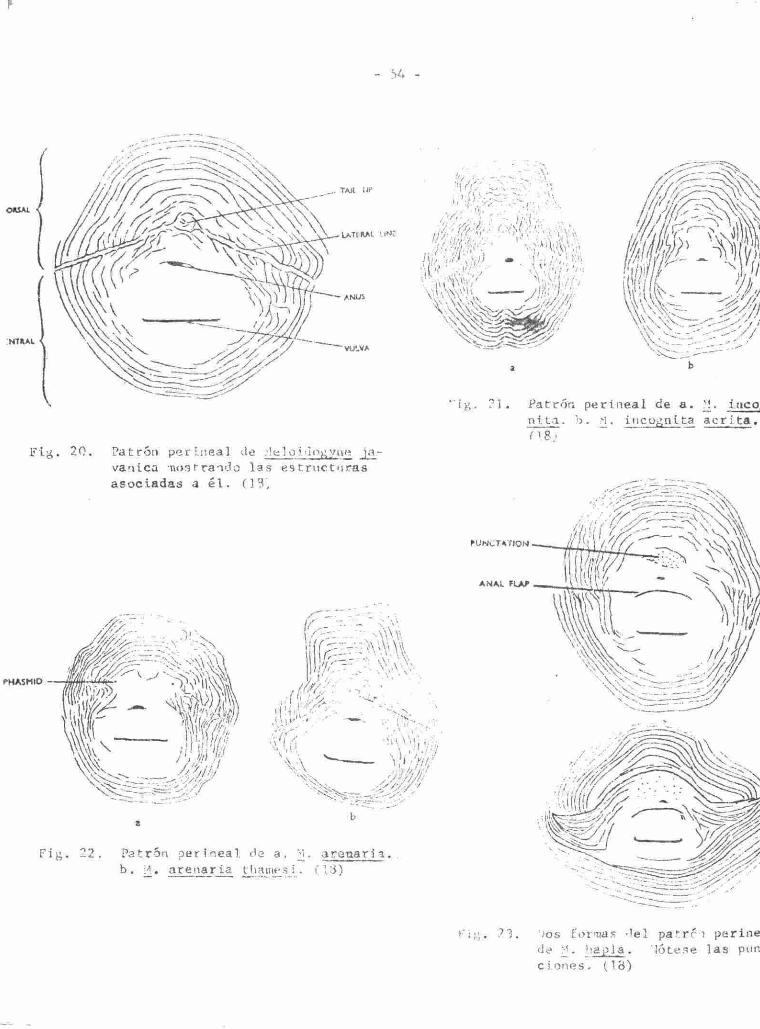
54
OMM.
INTItAL
_ . TAJl VIP
LATERAL LiNE
~ ANUS
F i g . 20 . Patrón p e r l n e a l de .•Itiloi Jii^yae ja -vanica mostrando l a s e s t r u c t u r a s a soc iadas a é l . (13)
PHASHIO
y' \\\\
mym m
^y^^\
,(My':^y -• ^ ' ^ , '
W'í ^$^sSÍ#f
yy y
• y y \ l ' í :
^%. . '\}>'yy 40
Fig. Patrón per inea l de a. M. a r e n a r i a . '̂ • il • a r e n a r i a t ha mes i . f 13)
% c ^ ^ .
ig . ?] . Patrón p e r l n e a l de a . M.. incog n t t a . b . M, incógni ta a c r i t a . f l8)
rUNCTATION
ANM. FUM"
i;.',. 73 . 'Jos Cormas ' lel patrc-i per iné da M. hap la . lótese las pun c iones . (18)

- 55 -
Se ha observado que la forma del patrón perlneal es bastante característica
para cada especie aunque es modificable por un biien número de factores. Pero
a pesar de esa variabilidad, Dropkln (15) opina que la configuración del pa
trón perlneal es controlada por factores genéticos.
Taylor, Dropkln y Martin (41) fueron los primeros en llevar a cabo un trabajo
sistemático usando ese parámetro. Las descripciones que siguen a continuación
son tomadas de su excelente estudio y de la obra de Thome (42).
** Meloldogyne javanlca;
Las líneas laterales son bien diferenciadas. Estrías de ambos sectores poco
o no interrumpidas. El arco generalmente es bajo y redondo, pero a veces al
to y estrecho. Estrías ventrales lisas. Los machos son escasos y diórquicos.
Ocurre en abundancia el Intersexo.
, Meloldogyne hapla;
Posee puntuaciones en el terminal del cuerpo, justamente por encima del afio.
Arco bajo, algo redondeado. Estrías de anibos sectores lisas o ligeramente on
duladas. Machos con uno o dos testículos. Ciertas colonias frecuentemente
no poseen machos, mientras que en otras, son numerosos. Cfi^urft)23).
^ Meloldogyne incógnita:
El patrón perlneal es algo oval y con arco irregular compuesto de estrías muy
juntas y onduladas. Las estrías de la parte interna del arco son numerosas,
en forma de zig-zag y quebradas. Los machos son mono o diórquicos.
Meloldogyne incógnita acrita:
Se diferencia de M. incógnita por poseer el arco más alto y mejor conformado,
y las estrías más separadas. Sin embargo, las estrías internas son más bien

56 -
ondula.da8. Los machos son monórqulcos o diórquicos.
Meloldogyne arenarla;
Las estrías son numerosas, cortas y desordenadas. El arco es más bien bajo
y redondeado. El patrón perlneal de M. arenarla se parece al de M.hapla.
Los machos son diórquicos.
Meloldogyne exigua:
El arco es bajo y ligeramente aplastado. Las líneas laterales son débilmen
te definidas.
Meloldogyne brevicauda:
El arco es ancho, alto. El patrón es rectangular y con estrías bien espacia
das. La cola del macho es corta y redonda y de allí deriva la denominación
brevicauda.
Meioidogyne tnornata:
ES parecido a M. incógnita, de la cual se diferencia por las estriaclones de
ia reglón labial del macho (1 en M. Inornata y 3 en M. incógnita).
CLAVE PARA ALGUNAS ESPECIES DE MELOIDOGYNE
(Tomada de Taylor, Dropkln y Martin (41) )
1. Arco bajo y redondeado o aplastado dorsalmente --------— 2
Arco alto, patrón oval o rectangular - - • r — - — 6
2. Líneas laterales bien diferenciadas y unidas en la punta
del patrón M. javanJca
Líneas laterales no diíerenciadas 3

- 57
3. Arco más o menos redondeado ---------a • 4
Arco achatado —---- 5
4. Fi~ecu£ntemente con puntuaciones por encima del ano M. hapla
Nunca con pHntnaaioaee jj, arenaria
5. Estrías plegadas cerca a laa líneas laterales M. exigua
Muchas estrías cortas cerca a las líneas laterales — H. arenaria thamesi
6. Patrón oval -- — ——---- 7
Patrón rectangular M̂. brevicauda
7. Arco mal formado, estrías dorsales juntas, onduladas o en zlg-sag -—
M. incógnita
Arco bien formado, estrías dorsales lisas a onduladas — — - - - - - — - —
_ . --_..—. .-.»._-.— M. incógnita acrita
Otra herramienta que ha sido muy empleada en la identificación de especies es
el esquema de Sasser (35), el cual está basado en las reacciones de las plan
tas en pruebas de patogenlcidad. De sus resultados, concluyó que M. hapla era
inmune a los cereales, al igual que lo eran M. incógnita. M. incógnita acrita
y posiblemente M. lavanica al maní. Así mismo se demostró que H. hapla era la
única especie que se podía identificar por el tipo de agallas, por ser peque
fias. Adeaáa M. hapla induce una proliferación de raíces, las cuales son den
sas en el sitio de infección severo. Su planteamiento queda sintetizado en
el esquema que sigue (35).

58 -
Población desconocida
Maní
Pepino.
\
\ L. peruvianua
Melón, trigo, avena o maíz
\ \
M. lavanica /
M. incógnita M. incógnita acrita M. hapla M. arena-rXi.
Identificación de especies de Meloldogyne por la reacción de las plantas (35).
Muchos investigadores han proseguido este tipo de ensayos. Townshend y David
son (45) encontraron que M. hapla parásita 41 especies de malezas que pertene
cen a las familias Compoaltae, Cruclferae, Polygonaceae y Caryophyllaceae.
Good, Minton y Jaworski (21) determinaron la eficiencia de Crot?)liria y de
Tagetes para contrarrestar el avance de varias especies de Meloidop.yne. En
contraron que C. spectabllis tenía un alto grado de resistencia contra M. ja
vanlca, M. incógnita. M. Incógnita acrita, M. arenaria y M. hapla. mientras
que Tagetes era altamente susceptible a M. aronatla y en menor grado a M. ha
pla.
Toledo y Davlde (43) estudiaron la patogenieidad de M. lavanica con bare en
la formación de agallas. Encontraron que a esta especie son susceptibles el
tabaco, tomate, cowpea, lechuga, pepino y coliflor, mientras que son resisten

- 59 -
tes el algodón, maíz, soya, trigo, maní, arroz, remolacha, papa y<cebolla.
De acuerdo con Fassullotls (17), todas las especies de Solanum melongena son
susceptibles a M. incógnita, mientras que la silvestre S. sisymbriifollum es
resistente. A este mismo método, es resistente Lycopersicon plmplnellifolium
y moderadamente susceptible L. peruvlanum (24).
Existen algunas especies de Meloldog'yne que atacan pastos y gramíneas. Rade-
vald y Pyeat (30) observaron que M, naasl se reprodujo muy bien sobre 27 gra
míneas y posteriormente Radewald et al. (29) ampliaron la gama de hospederos de
este nemátodo a cebolla, remolacha, alfalfa, algodón y Lycopersicon peryvlanum.
Sin embargo, encontraron que no se reproducía bien el maíz, Sorghum sudanenae
y Agrotis alba. Michell y Edwards (26) observaron que M. naasl era capaz de re
producirse sobre soya, aunque las poblaciones para ser patógenas deben ser muy
altas.
Otras especies de Meloldogyne que atacan gramíneas son M. graminls y M. grami-
nlcola (6, 20). Golden y Birchfleld (19) han registrado ataques de M. gramini-
cola en arroz.
A pesar de que las agallas radicales de Meloldogyne son las más comunes. Millar
y DiEduardo (27) reportaron un caso de una infección natural, sobrevivencia y
multipllcaaión de Meloldogyne Incógnita en hojas de Siderasis (Tradescantia)
fuácata. En las hojas, se encontraron h^nbras adultas, huevos viables y lar
vas mientras que las raíces estaban sanas. Hubo evidencia de que el nemátodo
entró por la yema follar.
6. GENERO HYPSOPERINE
El género Hypsoperine es parecido a Meloldogyne: sus características principa

60 -
0
.Vy-
^mm^̂ Fig. 24. Izquierda: Dibujos de hembras de lypsoperiae graminls parte anterior de un
espécimen. Derecha: Dibujos de machos y de larva del mismo nemátodo. A y B. Partes anterior y posterior de un macho. C, D y Ü. Partes anterior y po.st^ ri(-)r de una larva. (37)

61
loo oatÁm» o»a«rn,iAA d ol oii.idro No. IO. Sc descríbló por vez primera en 1964
(37) y hasta ei momento se conocen muy pocas especies (6,14,32,37). H. grami
nls. por ejemplo, es una asperje dlploide (n^ld) que se reproduce por parteno
génesis melótlca y probablemente por fertilización cruzada (45).
El género Hypsoperine ataca solamente monocotiledóneas pero no ataca maíz, to
mate, y zanahoria (37). La planta hospedante típica es Hordeum vulgare. We
bber y Barker (50) obser̂ 'aron que, a diferencia de Hypsoperine graminls. H.
ottersonl no parásita Agrostís pdlustrls.
Según Webber y Barker (50), las larvas de H. ottersonl emigran intercelular-
mente a través de la cortesa hacia la punta de la raíz, introducen su cabeza
en los tejidos localizados inmediatamente después del merlstema apical y lue
go se desarrollan paralelamente al eje longitudinal de la raíz. Las agallas
causadas por este nemátodo son bastante alimonadas o en forma de hoz y las cé
lulas gigantes se forman en las células no diferenciadas o patcialmente dife
renciadas que circundan a la cabeza del nemátodo en el cilindro vascular.
7. GENERO MELOTDODERA
Meloldodera es muy parecido al grupo Heterodera rostochiensis y a algunas es
pecies de Meloldogyne. Sus característiéas más sobresalientes están seflaladas
en el cuadro No. 11. Entre las pocas especies conocidas, se halla M. floriden-
sls, la cual ae reproduce por partenogénesis mltótlca (2n-26). Este género a-
taca más bien árboles forestales.
8 . GEÍÑERO CRYPH0D3RA
Este género parece estar más relacionados con Meloldodera que con los demás

62 -
' ' ' i ¿ . 2 5 . '• 'áloido.'o>-ra _f b r r i .li:ns i .•. . A. Cabe'iT ÍÍP i!!i,3 l a r v a , vi.tit a l a t e r a l . B. c abeza de I.4 o.j-i l ) r a . C.. Pat. r o n , p e r i n e . a l , L'. Co l a de la l a r v a , v i s t a l a t e r a l , ii. Regióa de l es6fa ;40 . c. Pat r ó n v u l v a l . (Toma.lo de i ' a i , 0 6 . C i t . en e l
c a p f t ' i l o .^n t í • , r i o r ) .

- 63
g£n«ro0 de la fantil in Heteroderidae. Las hembraA Bon o v a l e s , de cutícula eape
sa y no ponen los huevos en matriz. No poseen un quiste tantresistente como
en Heterodera. La especie típica C. euealypti se encontró alrededor de las ra
ices de Eucalyptus malor y es capaz de reproducirse sobre otras especies de eu
caliptos. Se distingue de Meloldodera florldensls. M. clares y M. armenlaca
por tener la vulva mucho más posterior al ano (45-70 mieras.).

^PS^n ss ^ f ^ ' '.•
- 64 -
Fig. 26. Cryphodera eacalyptl. Macho. A. Vista lateral. 3. Cabeza. C. Vista de freate. D. Sección tran.sversal en la rcilón de los campos laterales. £. Campos latera les en la mitad del cuerpo. F. Vista lateral de la cola. G. Vista ventral de la cola. (10)
i^Mi

65 -
tiiiversid.id Mücional de Culn'^rinia DfparUKr.n'u df Piil.lii'Vii:''ti^'í
Medellin
y.-y'-yy::Uy^;^r
'^mymy / • —' y^ryi^y'iípy
i ^ \ yfftí
*y-y'><''^^0^>^iyé • • "'""--. •
m m ^ ^ y W :• yt-... .
0$ f • ^y '^y:>n^0^i •• y - ' y y y y ' - ^ ! y y y '̂ v y ^ ' y y a > y •
AC'Toir
B
Fig, 27. Cryphodera encalypt lateral. H. Cabeza, de la vulva. (10)
30)i.
Hembra . A. Vista E s t r í a s a l r ededor
Fig. 28. Cryphodera encalypt i. Segundo e tado larvario. A. Vista lateral. B. Cabeza. C, Vista de frente. D. Sección transversal del campo lateral en la mitad del cuerpo. E. Vista ventral de la región an. F. Cola. (10)

- 66
Cuadro No TO. CARACTERÍSTICAS DE ALGUNOS GÉNEROS DE LA FAMILIA HETERODERIDAE
MACHOS : Cabeza
Anfldas
Estilete
Testículo
Espíenla
HQIBRA :
Cuerpo
Cabeza
Estilete
Poro excretor
M E L O I D O G Y N E
Región del labio con o sin ánulos. Número variable. Rodeada por capa anterior
Labios laterales más grandes que los submedlanos.
Sacos anfIdiales conspicuos
Bien desarrollado. Largo Inferior a 24 mieras.
Uno o dos
Casi terminal
INO forma quiste p í-
Forma de pera. Permanece blando y relativamente blanco.
Labios laterales mas grandes ¡que los submedlanos. Masa cefálica esclerotizada.
Delgado con bulbos débilmente desarrollados. 13-16 mieras de largo.
No muy anterior al metacorpus 12-27 ánulos después de la ca beza.
H E T E R O D E R A
Región del labio anulada. Número vlarlable. .lusoncia de capa anterior.
Labios laterales Iguales o más pequeños que los submedlanos.
Sacos anfldlales pequeños.
Bien desarrollado. Largo superior a 24 mieras.
Uno
Casi terminal
Forga quiste
Esférico,de limón o de pera. Se endurece y se oscurece frecuentemente.
Labios no son fácilmente di¿ tlngulbles. No hay esclerotlzaclón.
Muy delgado con pequefios bujl bos. 27 mieras de latgo.
Posterior al metacoppus.Apro ximadamente al nivel del anl lio nervioso.
H Y P S O P E R I N E
Anulaciones no diferenciadas
Corto. Largo de 18 mieras
Uno
Casi terminal
No forma quiste
Oval - blando
Esclerotlzaclón no dife rendada. ""
Desarrollado. 12-13 mi €ras de largo.
Al nivel de los bulbos del estilete o ligeramente posterior a ellos.
M E L 3 I D O D E R A
Anulada
Tamafio intermedio
Corto
Corto
No forma quiste
Pera, espeso, blan ca.
Posterior al -metacorpus .

- 67 -
- 2 -
Anulación
Huevos
Glándulas rectales
Larva en 2o. Estado
Estilete
Cola
Larva en 8o. Estado Cola
PARASITISMO
N O T A :
M E L O I D O G Y N E
Transversal. Circular en la re glón anal. Uso en la clasificación.
Depositados en matriz
6
Región labial bien definida. Masa cefálica esclerotizada. Con D-4 ánulos.
Delgado. Bulbos bien definí. dos. 10 mieras de largo.
Terminal agudo
Las larvas siguen teniendo cola puntiaguda hasta la última muda.
Formación de agallas.
Generalmente las hembras son endoparásitas.
H E T E R O D E R A
No transversal
Frecuentemente retenjL dos en el cuerpo.
Ánulos variables. Masa cefálica muy escle rotizada.
Bien desarrollado, largo de 20-25 mieras
Terminal agudo
No puntiaguda
No formación de agallas.
Hembras semlendopará sitas. - *"
H Y P S O P E R I N E
Estrías transversales no bien diferenciadas en la parte po¿ terior del cuerpo. Uso en la clasificación.
Generalmente depositados en ma triz.
6
Anulaciones no visibles
Terminal redondo, no agudo
Agallas pequefias
Sembras endoparásitas
M E L O I J O D E R A
Uso en la clasificación.
Retenidos en el cuerpo.
Muy esclerotizadas. Ánulos muy pronunciados.
Terminal agudo
No formación de aballas.
Hembras semiendoparásitas
1. 2.
La raya Indica que los datos son aún desconocidos. El cuadro fué elaborado y ampliado a partir de la muestra original aparecida en el articulo ^^ Franklln (18).

. 68 -
B I B L 1 0 G R A ^ I A
1. ALLEN, M.W. y S.A, Sher. 1967. Taxonootlc probjem* conceming the phy-
toparasltlc nematodes. Ann. ftev. Phy. 5:247-264.
2. ALLEN, M.W. 1952. Observation^ on the genus Meloldogyne Goeldi, 1887.
Proc. Helv. Soc. Wash. 19(l):44-51.
3. BADALKJODZHAYEV, I. 1971. Embrryogenesls of Meloldogyne lavanica (Ty
lenchlda, Heteroderidae). Zool. Zh. 50(11): 1621. En Tob. Absts.
16(3):283. 1972.
4. BALASUBRAMANION, M. y G. RANGASWAMI. 1963. On ogg number and size pro
duced by Meloldogyne javanlca. the root-knot nematode, on five host
plants. Bangalore. Cur. Sel. 32(12): 562-563. En Tob. Absts. 9
(l):86-87. 1965.
5. BALASUBRAMANIAN, M. y G. RANGASWAMI. 1964. Studies on host range and
histopathology of root-knot Infectlon caused by Meloldogyne javanlca.
Ind. Phy. 17(2):126-132.
6. BIRCHFIELD, W. 1965. Host-paraslte relatlons and host range studies
of a new Meloldogyne specles ln southem USA. Phy 55:1359-1361.
7. BIRCHFIELD, W. 1964. Histopathology of nematode - Induced gall of
Echinoclúa colonum. Phytopathoiogy 54:888.
8. BIRD, A.F. y G.E. ROGERS. 1965. Ultrastructure of the cutlcle and Its
formatlon ln Meloldogyne Javanlca. Nematologica 11(2):224-230. En
Tob Absts 10(1): 1966.
9. BIRD, A.F. y G.E. ROGERS. 1965. Ultrastructure and histochemical
studies of the cells producing the gelatlnous matrix ln Meloldogyne.
Nematologica 11(2): 231-238. En Tob Absts 10(l)t58-1966.

- 69 -
10. COLBRAN, R.C. 1966. Studies of plant and soil nematodes. 12T The
eucalypt cystold nematode Gryphodera euealypti, n. sp., (Nematoda:
Heteroderidae) a paraslte of eucalypts ln Queensland.Queen. Jour.
Agrie. Anim. Sel. 23(l):41-47.
11. DAVIDE, R.G. y A.C. TRIANTAPHYLLOU. 1965. Influence of the evlron-
ment, on the development and sex differentlatlon of root-knot nemato
des. II Effect of foliar apllcatlon of malelc hydrazlde. Nematologi
ca 14(l):37-46. En Tob. Absts. 12(7-8): 452. 1968.
12. DAVIDE, R.G. y A.C. TRIANTAPHYLLOU. 1965. Influence of foliar apllca-
tions of maleic hldrazlde on varlons host plants repon sex differen
tlatlon in Meloldogyne javanlca and Meloldogyne incógnita. Nematolo
gica 11(1): 35. En Tob. Absts. 9(10):554. 1965.
13. DE A. SANTOS, M.S.N. 1972. Productlon of male Meloldoc¡vne and atraction
to their famales. Nematologica 18(3):291-302. En Tob. Absts. 17(4):
332. 1973.
14. DICKERSON, O.J. 1966. Some observations on Hypsoperine graminls in kan-
sas. Plant Dis. Rept. 50(6):396-399.
15. DROPKIN, V.H. 1963. Studies on the variabllity of anal píate pattems
in puré Unes of Meloldogyne spp. The root - knot nematode Proc.
Helm. Soc« Wash. 20(1):32-39.
16. ELSEA, J.R. 1951. The hlstological anatomy of the nematode Meloldogyne
hapla. (Heteroderlda). Proc. Helm. Soc. Wash. 18(1):53-63.
17. FASSULIOTIS, G. 1973. Susceptiblllty of eggplant, Solanum m̂ ilongena. to
root-knot nematode, Meloldogyne incógnita. Plant. Dis. Rept 57(7):
606-608.

"^
- 70 -
18. FRANKLIN, M.T. 1^60. loot-Knot nematodes, Meloldogyne spp. En Plant
Nematology. ed. Southey, J.F. Section 11:49-61. Mlnlstry of agrlcul
ture, Fisherles and Food. Technlcal Bulletln No. 7 London 175 p.
19. GOLDEN, A.M. y W. Birchfleld: 1968, Rice root-knot nematode (Meloldogyne
gvamlnleola) as a new pest of rice. Plant. Dis. Rept. 52(6):423.
20. GOLDEN, A.M. y W. BIRCHFIELD. 1965. Meloldogyne granAnlcola (Heterode-
tidae) a new specles of root-knot nematpide from grass. Prod. Helm.
Soc. Wash. 32(2):223-231.
21. GOOD, J.M., N.A. MINTOII y C.A. JAWORSKI. 1965. Relativa susceptiblll
ty of selected cover crops and coastal bermuda grass to plant nemato
des. Phy 55(9)1026-1030.
22. GRIFFIN, G.D. y E.C. JORGENSON. 1969. Life cycle and reproductlve of
Meloldogyne hapla on potato. Plant Dis Rept. 53(4):259-262.
23. JONES, M.G. y D.A. N0RTHC0T2. 1972. Multlnucleate transfer cells indu
ced ln coleus roots by the root-knot nematode, Meloldogyne arenaria.
Protoplasma 75(4):381-395. En Tob. Absts 17(2):131.
24. KHAN, A.M., S.K. SAXENA y Z.A. SIDDIQI. 1971. Reaction of some culti-
vated and Wild specias of Lycopersicon and cucúrbita to Maloldogyne
incógnita (Kofoid y White) Chltowood. Ind. Phy 24(1):197-200i
25. MAGGENTI, A.R. y M.W. ALLEN, 196Q. The prlgin of the gelatlnou matrix
ln Meloldogyne. Proc. Helm. Soc. Wash. 27(1);4-10.
26. MICHELL, R.E. y D.l. EDWARDS. 1973. Susceptiblllty of soybean to Melol
dogyne naasl. Plant Dis. Rept. 57(3):207-209.
27. MILLER, H.N. y A.A. DIEDUARDO. 1962, Leaf galls on Siderasis fuscata
caused by root-knot nematode Meloldogyne incógnita. Phy 52(1):22

71
28. OWENS, R.G. y H.M. NOVOTNY. 1969. Physiologicaí and biochemical stu
dies on liematode galls. Phy 50:650
29. RDDEMALD, J.D. et al. 1970. Meloldogyne naasl. a paraslte of turfgrass
in southen California. Plant Dis. Respt 54(11):940-942.
30 MDEWALD, J.D. y L.E. PYEATT. 1966. Ocurrence, host range and control
studies for Meloldogyne naasl on turf. Phy 56:896.
31. RANGAS-WAMI, G. y M. BALASUBRAMANIAN. 1964. Presence of aminoaclds and
Índole compounds in the root-knot affected plants. Ind. Phy 17:234-
237.
32. RAN, G.J. y G. FASSULIOTIS. 1965. Hypsoperine epartinae; a gall-forming
nematode on the roots of smooth cordgrass. Proc. Helm. Soc. Wash.
32(2):159-162.
33. RIFFLE, J.W. 1963. Meloldogyne ovalls (Nematoda: Heteroderidae) a new
specles of toot-knot nematode. Proc. Helm. Soc. Wash. 30(2):287-292.
34. ROHDE, R.A. 1972. Expresslon of resistance ln plants to nematodes Ann.
Rev. Phy. 10:233-253..
35. SASSER, J.N. 1952. Identification of rootsknot nematodes (Meloldogyne sp)
by host reactions. Plant Dis. Rept. 36(3):84-86.
36. SETTY, K.B.H. y A.W. WHEELER. 1968. Growth substances ln roots of toma-
to (Lycopersicon esculentum Mili. ) infected with roof-k-hot nematodes
(Meloldogyne spp.) Ann. appl. Biol. 61(3):495-501.
37. SLEDOS, E.B. y A.M. GOLDEN. 1964. Hypstoperine graminls (Nenatoda Hete-
roderldae), a new genus and specles of plant-parasltic nematode. Proc.
Helm. Soc. Wash. 31(1):83-88.
38. SLEDGE, E.B. 1962. Preliminary respect on Meloldogyne sp. paraslte of
grass ln Florida. Plant Dis. Rept. 46(1):52-53.

72 -
39. SIDDQüI, I.A. y D.P. TAYLOR. 1970. Symptoms and varletal reaction. of
oats to the Illinois of the barley root-knot nematode, Meloldog'yne
naasl. Plant. Dis. Rept. 54(11):972-975»
40. STOYANOV, D. 1967. Especies de nemátodos parásitos del plátano en Cuba
y posibilidades de control. Rev. Agrie. (Cuba) l(3):9-47.
41. TAYLOR, A., V.H. DROPKIN y Q.C. MARTIN* 1955. Perlneal pattems of
root-knot nematodes, Phy 45(1):26-34.
42. THORKE, G. 1961. Prlucipiássof nematology. McGraw-Hlll Book Company,
Inc. 551 p.
43. TOLEDO, R.T. y R.G. DAVIDE. 1969. 1969. Reactions of dlfferent vege
table and fleld cropo to infectlon by Meloldogyne lavanica. Philipp.
Phy. 5(l/2):45-54. En Tob. Absts. 15(5):511. 1971.
44. TOWNSHEND, J.L. y T.R. DAVIDSCN. 1962. Some weed hosts of the northen
root-knot nematodes Meloldogyne hapla Chitwood, 1949 ln Ontario. Can
Jour. Bot. 40(4):543-547.
45. TRIANTAPHYLLOU, A.C. 1969. Cytogenetlc evoluation of the phylogeny in
the famlly Heteroderidae. En Proceedlngs of the Symposlum on Tropi
cal Nematology. Ed. Ramos, J.A. et al. 11-17. Agrie. Exp, Sta. Puer
to Rico. 169 p.
46. TRIANTAPHYLLOU, A.C. 1966. Polyplldy and reproductlve pattems in the
toot-knot nematode Meloldogyne hapla. J. Morphol. 118(3):403-414.
En Tob. Absts 17(4): 725. 1966.
47. TRUDGILL, D.L. 1972. Influence of feedlng duratlon on moulting and
sex determination of Mnlotdo^yne incógnita. Nematologica 18(4):
476-481. En Tob. Absts 17(4):339. 1973.

73
48. VAGMA, M.K. et al. 1971. Male intersexes ln Meloldogyne Incógnita from
India. N^natological 17(3):49l. En Tob. Absts. 16(2):170. 1972.
49. VIBLIERCHEIO, D.R. 1971. Nematodes and other pathogens ln auxin-rela-
ted plant growth dlsorders. Bot Rev. 37(1):1-21.
50. WEBBER, A.J. y R.R. BARKER. 1967. Biology of the pseudo root-knot nema
tode Hypeoperlne graminls. Phy. 57:723-728.
51. Wouts, W.M. 1972. Revisión of the famlly Heteroderidae (Nematoda:Tylen
choldea)Nematologica 18(4):439-446. En Tob. Absts: 17(4):341. 1973
52. WOUTS, W.M. y S.A. SHER. 1971. The genera of the subfamlly Heteroderi
nae (Nematoda:Tylencholdea) with a descrlptlon of two new genera.
Journal of Nematology 3(2):129-137.